
Download
ORIGINAL ARTICLE
RNF20/RNF40 ameliorates streptozotocin-induced type 1 diabetes by activating vitamin D receptors in vivo
Junxiang Liu*, Xuri Wu, Haixia Qin, Ying Hu, Zhiyun Zhang, Yanmei Wang, Jinlan Li
Department of Endocrinology, Ordos Central Hospital, Ordos, Inner Mongolia Autonomous Region, China
Abstract
Background: Type 1 diabetes is one of the chronic autoimmune diseases. Its features include the immune-triggered pancreatic beta-cells destruction. Ubiquitin ligases RNF20 and RNF40 have been discovered to participate into beta cells gene expression, insulin secretion, and expression of vitamin D receptors (VDRs). However, no reports about the role of RNF20/RNF40 in type 1 diabetes are known till now. The aim of this study was to clarify the role of RNF20/RNF40 in type 1 diabetes and explore the mechanism.
Methods: In this study, streptozotocin (STZ)-induced mice type 1 diabetes model was used. The protein expressions of genes were examined through Western blot analysis. Fasting blood glucose was detected through glucose meter. The plasma insulin was tested through the commercial kit. Hematoxylin and eosin staining was utilized to observe pathological changes of pancreatic tissues. Immunofluorescence assay was performed to evaluate the level of insulin. The levels of pro-inflammatory cytokines in serum were assessed by enzyme-linked-immunosorbent serologic assay. The cell apoptosis was measured through terminal deoxynucleotidyl transferase dUTP nick end labelling assay.
Results: STZ was used to stimulate mice model for type 1 diabetes. At first, both RNF20 and RNF40 expressions were down-regulated in STZ-mediated type 1 diabetes. Additionally, RNF20/RNF40 improved hyperglycemia in STZ-stimulated mice. Moreover, RNF20/RNF40 relieved pancreatic tissue injury in STZ-induced mice. Further experiments found that RNF20/RNF40 rescued the strengthened inflammation mediated by STZ treatment. The cell apoptosis was enhanced in the pancreatic tissues of STZ-triggered mice, but this effect was weakened by overexpression of RNF20/RNF40. Besides, the VDR expression was positively regulated by RNF20/RNF40. Finally, VDR knockdown reversed improved hyperglycemia, inflammation, and cell apoptosis stimulated by overexpression of RNF20/RNF40.
Conclusion: Our findings proved that RNF20/RNF40 activated VDR to relieve type 1 diabetes. This work might highlight the functioning of RNF20/RNF40 in the treatment of type 1 diabetes.
Key words: RNF20/RNF40, STZ, type 1 diabetes, VDR
*Corresponding author: Junxiang Liu, Department of Endocrinology, Ordos Central Hospital, No. 23, Yijin Horuo West Street, Dongsheng District, Ordos, Inner Mongolia Autonomous Region 017000, China. Email address: [email protected]
Received 8 December 2022; Accepted 9 March 2023; Available online 1 July 2023
Copyright: Liu J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Type 1 diabetes (also called insulin-dependent diabetes mellitus) is one of the chronic autoimmune diseases. It is caused by interaction between environmental triggers and individual genetic factors.1,2 Type 1 diabetes is discovered in all age groups, and its main features include unusual autoimmune destruction of islet beta cells, resulting in complete insufficiency of insulin synthesis.3–5 This disease forces patients with type 1 diabetes to undergo lifelong hormone replacement therapy.6,7 Therefore, more studies are conducted to explore novel molecular markers for the treatment of type 1 diabetes.
Type 1 diabetes and insulin-resistant individuals are linked with the deficiency of vitamin D.8 In addition, it is discovered that a simultaneous increase in the prevalence of diabetes and the deficiency of vitamin D globally could be possibly due to inadequate exposure to sunlight to inhibit vitamin D synthesis in the skin and inadequate dietary supply.9 Vitamin D primarily functions by combining to vitamin D receptors (VDR), as two single nucleotide polymorphisms in VDR are linked with type 1 diabetes.10 These findings strongly imply the vital functions of the vitamin D/VDR axis in diabetes.
Ubiquitin ligase RNF20/RNF40 (also called as BRE1 complex) was first authenticated as an E3 ubiquitin ligase of histone H2B.11 RNF20 helps in tumor suppression by regulating certain genes and maintaining genomic stability.12,13 Research studies have demonstrated that lack of RNF20 or RNF40 affects the expression of key genes in islet beta cells.14 The RNF complex is indispensable for glucose-stimulated insulin secretion and normal levels of mitochondrial reactive oxygen species (ROS), demonstrating that RNF20 and RNF40 regulate expression of genes and insulin secretion in islet beta cells.15 Furthermore, it has been reported that the deletion of RNF20 or RNF40 reduces the messenger RNA (mRNA) and protein levels of VDR in inflammatory bowel disease, thereby aggravating intestinal inflammation.16 However, since the roles and related regulatory mechanisms of RNF20 and RNF40 in type 1 diabetes are still unclear, these are further investigated.
In this study, the functions and regulatory influences of RNF20 and RNF40 on the progression of type 1 diabetes are examined. Our results demonstrated that RNF20 and RNF40 relieved STZ-induced type 1 diabetes by activating VDR. This work could provide useful bio-targets in the treatment of type 1 diabetes.
Materials and Methods
Mice model
C57BL/6J male mice (6–8-week old, n = 50) were used in the present study (Vital River Laboratory Animal Technology Co. Ltd., Beijing, China). The mice were maintained in cages under a 12-h light/dark cycle with free water and food. All animal experiments were authorized by the Animal Care and Use Committee of Ordos Central Hospital, Ordos.
In this study, streptozotocin (STZ) induced mice type 1 diabetes model was used. Mice were intraperitoneal injected with 50-mg/kg STZ (Sigma, St. Louis, MO) in phosphate-buffered saline solution (PBS) once a day for 5 days to create type 1 diabetes model. The blood glucose level was examined, and 300 mg/dL or more was considered as diabetes. Mice were anesthetized with 2.5% isoflurane. Finally, mice were euthanized, and blood and pancreas were collected for further experiments. In addition, mice were sacrificed after 10 days following the STZ treatment.
The mice were randomly separated into the following groups: control, STZ, STZ+vector (the vector plasmid which did not overexpress any proteins), STZ+RNF20, STZ+RNF40, STZ+vector+sh-NC (negative control), STZ+RNF20+sh-NC, STZ+RNF40+sh-NC, STZ+RNF20+sh-VDR, and STZ+RNF40+sh-VDR (n = 5 in each group). Mice in STZ, STZ+vector, STZ+RNF20, STZ+RNF40, and STZ+vector+sh-NC groups were treated with STZ for 10 days. Mice in STZ+RNF20+sh-NC, STZ+RNF40+sh-NC, STZ+RNF20+sh-VDR, and STZ+RNF40+sh-VDR groups were treated with STZ and transfected with indicated plasmids. Plasmid and short hairpin RNA (shRNA, 10 µg) were transfected in vivo using transfection reagents (10 µL).
Western blot analysis
Proteins were extracted from pancreatic tissues and subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were moved onto polyvinylidene fluoride (PVDF; Beyotime, Shanghai, China) membranes. Non-fat milk (5%) was utilized for sealing the membranes; then the primary antibodies were supplemented onto the membranes for 12 h. Next, appropriate secondary antibodies (1:2000; ab7090; Abcam, Shanghai, China) were added for another 2 h. The blots were evaluated with chemiluminescence detection kit (Thermo Fisher Scientific Inc., MA). Blots were normalized to β-actin, and quantified with the Image J software (NIH, Bethesda, MD). Primary antibodies were anti-RNF20 (1 µg/mL, ab32629; Abcam), anti-RNF40 (1:1000, ab191309), anti-Bcl-2-associated X (Bax, 1:1000, ab32503), anti-cleaved caspase-3 (1:5000, ab214430), anti-cleaved-poly (ADP-ribose) polymerase (PARP, 1:1000, ab32064), and anti-β-actin (1 µg/mL; ab8226).
Detection of fasting blood glucose and plasma insulin
Blood glucose was determined using the FreeStyle Lite blood glucose meter (Abbot Diabetes Care Inc. California, USA). Plasma insulin level was measured by insulin EIA kit in line with the manufacturer’s instructions (ALPCO Diagnostics, Salem, NH).
Hematoxylin and eosin (H&E) staining
Pancreas were put in 4% paraformaldehyde and subjected to paraffin-embedding. Sections, 5-μm thick, were cut, and stained in H&E solution. Light microscope (Olympus Corporation, Tokyo, Japan) was used to observe prepared sections.
Immunofluorescence assay
Pancreatic tissues of the mice were kept in paraformaldehyde (4%; Sigma) and Triton X-100 (0.2%; Sigma). Post-washing, the tissues were sealed with bovine serum albumin (BSA 4%) overnight at 4°C. Next, the primary antibody of insulin (1:200, ab181547; Abcam) was supplemented overnight at 4°C. The secondary antibodies were added and incubated. The staining of nucleus was performed with 4',6-diamidino-2-phenylindole (DAPI), and observed using Fluorescence microscope (Olympus).
Terminal deoxynucleotidyl transferase dUTP nick end labelling (TUNEL) assay
The in situ cell death detection kit (Minneapolis, MN) was selected for TUNEL assay. Pancreatic tissue sections (5 μm) were deparaffinized and hydrated, and immersed in proteinase K solution. The sections were incubated with 3% H2O2. Post-rinsing, TUNEL solution (50 μL) was added for 1 h in dark. The staining of nucleus was done with DAPI. Fluorescence microscope (Olympus) was used to determine cell apoptosis.
Enzyme-linked-immunosorbent serologic assay (ELISA)
The levels of inflammation markers, such as interleukin (IL)-6, monocyte chemoattractant protein-1 (MCP-1), tumor necrosis factor-α (TNF-α), IL-1β, IL-8, and interferon gamma (IFN-γ), were inspected in mice serum. Pro-inflammatory cytokines in serum were measured using the corresponding kits (Abcam).
Statistical analysis
The GraphPad Prism software, version 8.0 (GraphPad Software, La Jolla, CA) was employed for analysis. The data were represented as mean ± standard error of mean (SEM). The analysis of comparisons between two or more groups was performed through the Student’s t-test or one-way analysis of variance (ANOVA). P < 0.05 was defined as statistically significant.
Results
RNF20/RNF40 improved hyperglycemia in STZ-stimulated mice
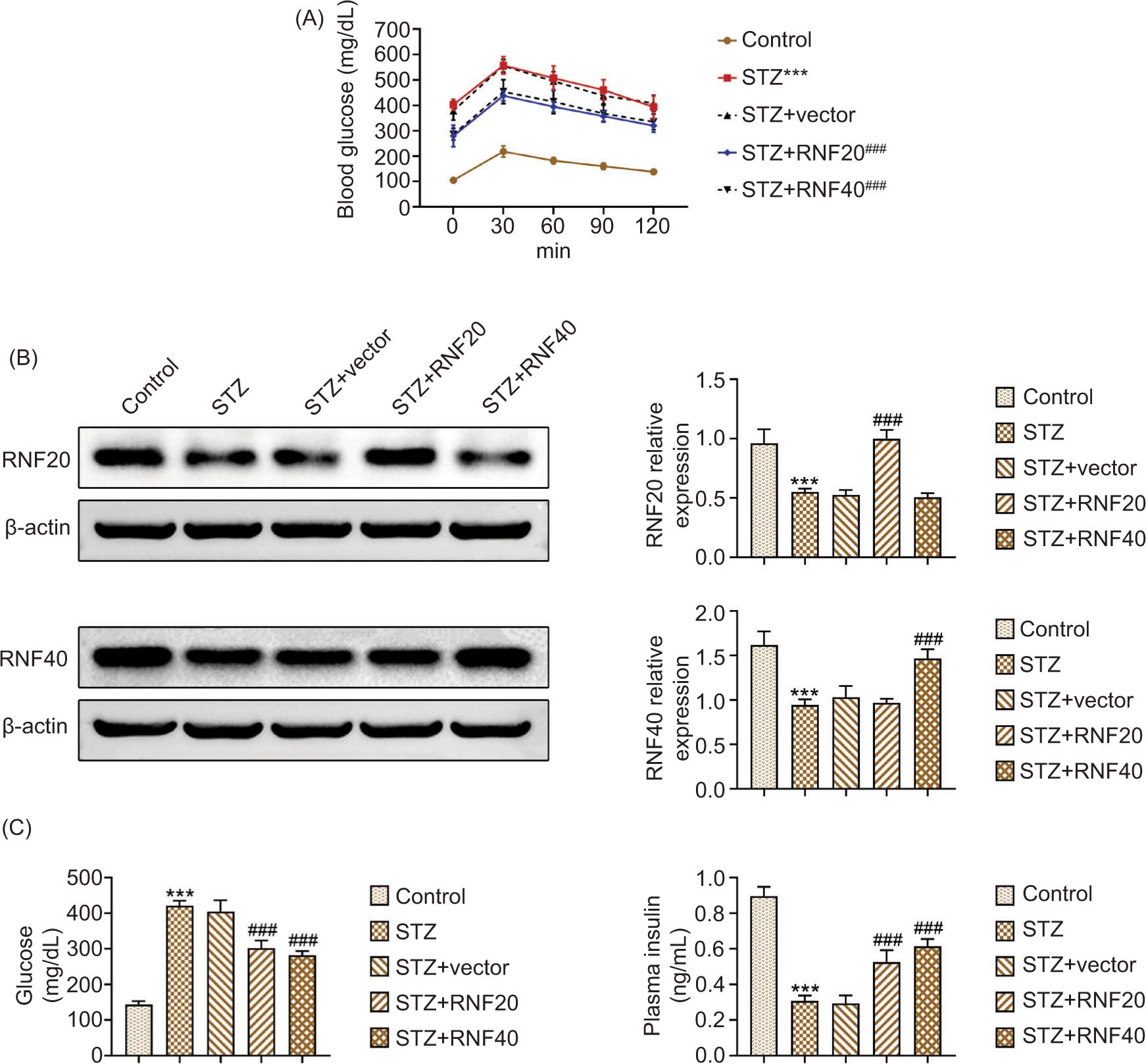
The blood glucose tolerance curve is shown in Figure 1A. As displayed in Figure 1B, the protein expressions of RNF20 and RNF40 decreased in STZ-induced mice, but their expressions increased after overexpression of RNF20 or RNF40. Next, as demonstrated, glucose increased and plasma insulin decreased in STZ-stimulated mice, but these effects were reversed after up-regulating RNF20 or RNF40 (Figure 1C). These results clarified that RNF20/RNF40 improved hyperglycemia in STZ-stimulated mice.
Figure 1 RNF20/RNF40 improved hyperglycemia in STZ-stimulated mice. Mice were divided into the following groups: Control, STZ, STZ+vector, STZ+RNF20, and STZ+RNF40, with 6 mice in each group. (A) Blood glucose was observed in the indicated groups at 0, 30, 60, 90, and 120 min. (B) The protein expressions of RNF20 and RNF40 were examined through Western blot analysis. (C) Fasting blood glucose level was measured using glucose meter, and plasma insulin was evaluated by commercial kit. ***P < 0.001 vs the control group; ###P < 0.001 vs the STZ+vector group.
RNF20/RNF 40 ameliorated pancreatic tissue injury in STZ-induced mice
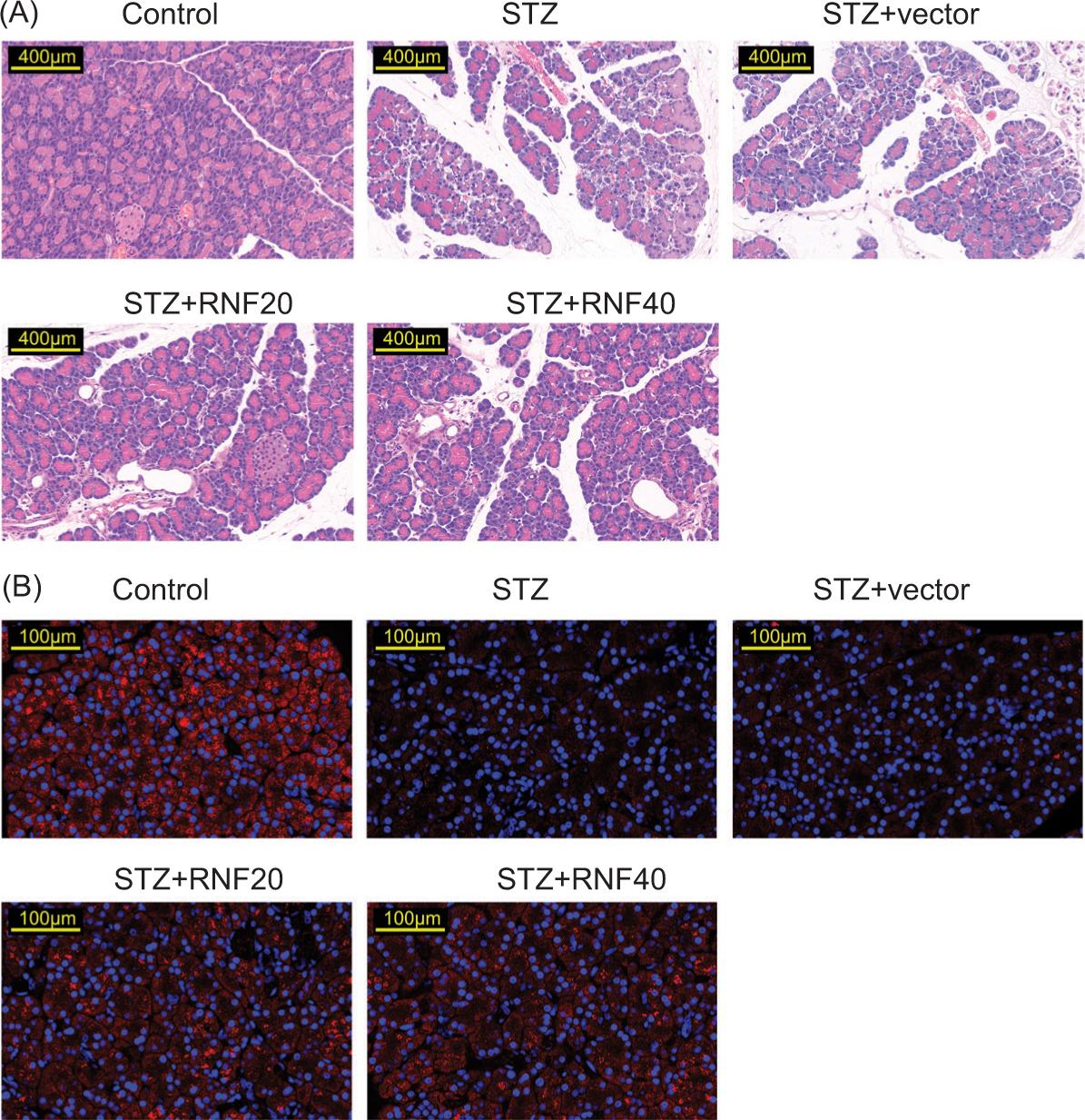
H&E staining discovered decrease in the morphology of the pancreas, reduction in the islets, and appearance of inflammatory cell infiltration; however, these changes ameliorated after RNF20 or RNF40 overexpression (Figure 2A). Additionally, the insulin level was reduced in STZ-mediated mice, but this effect was offset after the overexpression of RNF20 or RNF40 (Figure 2B). Taken together, RNF20/RNF40 ameliorated pancreatic tissue injury in STZ-induced mice.
Figure 2 RNF20/RNF40 ameliorated pancreatic tissue injury in STZ-induced mice. Mice were divided into the following groups: Control, STZ, STZ+vector, STZ+RNF20, and STZ+RNF40. (A) H&E staining was used to observe pathological changes in pancreatic tissues. (B) Immunofluorescence was performed to evaluate the level of insulin.
RNF20/RNF40 relieved inflammation in STZ-mediated mice
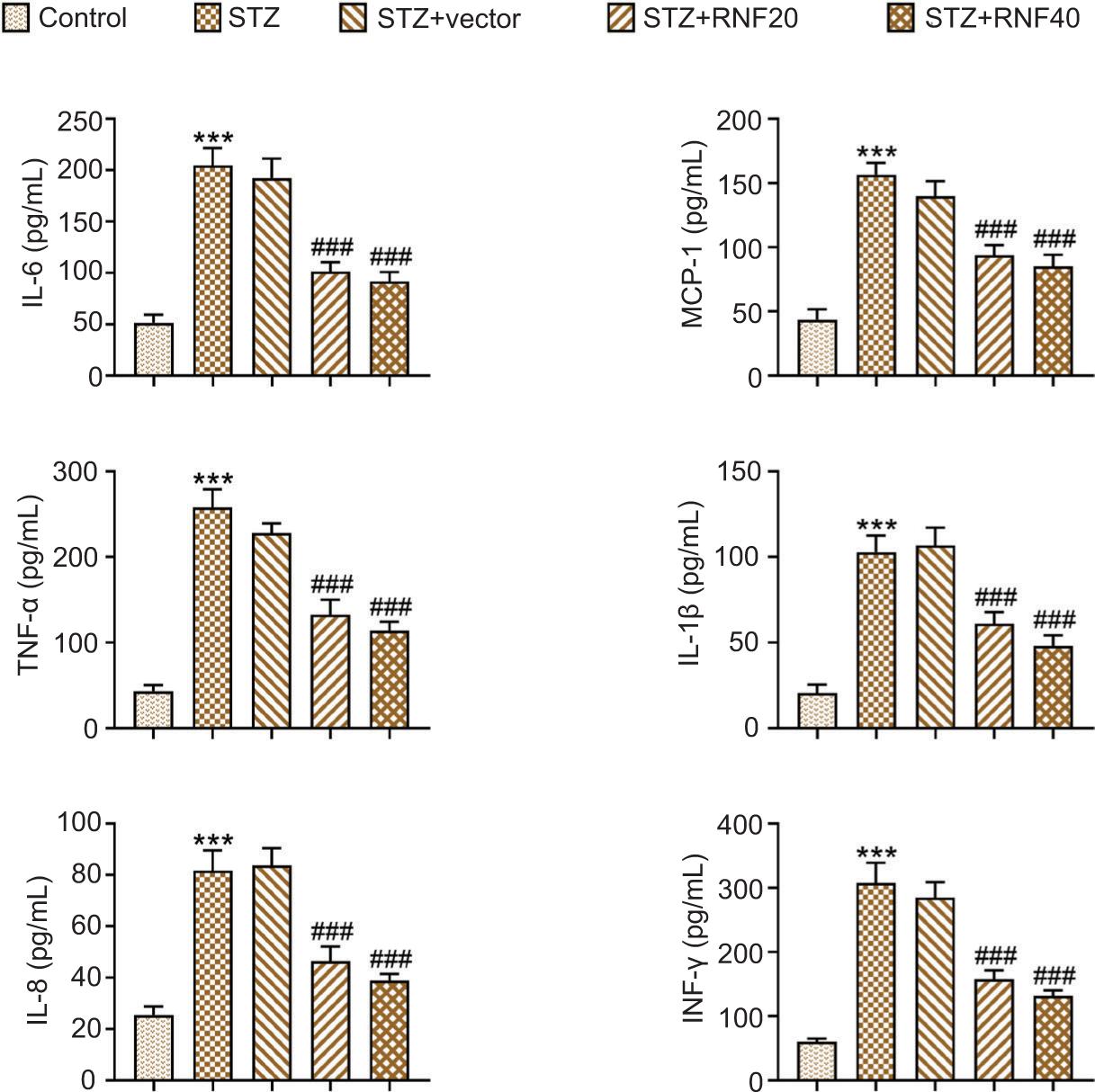
Furthermore, ELISA showed that the levels of IL-6, MCP-1, TNF-α, IL-1β, IL-8, and INF-γ (pro-inflammatory cytokines) were enhanced in STZ-induced mice, but these changes were attenuated after the overexpression of RNF20 or RNF40 (Figure 3). In general, RNF20/RNF40 relieved inflammation in STZ-mediated mice.
Figure 3 RNF20/RNF40 relieved inflammation in STZ-mediated mice. Mice were divided into the following groups: Control, STZ, STZ+vector, STZ+RNF20, and STZ+RNF40. The levels of IL-6, MCP-1, TNF-α, IL-1β, IL-8, and INF-γ in serum were assessed by ELISA. ***P < 0.001 vs the control group; ###P < 0.001 vs the STZ+vector group.
RNF20/RNF40 improved cell apoptosis in the pancreatic tissues of STZ-triggered mice
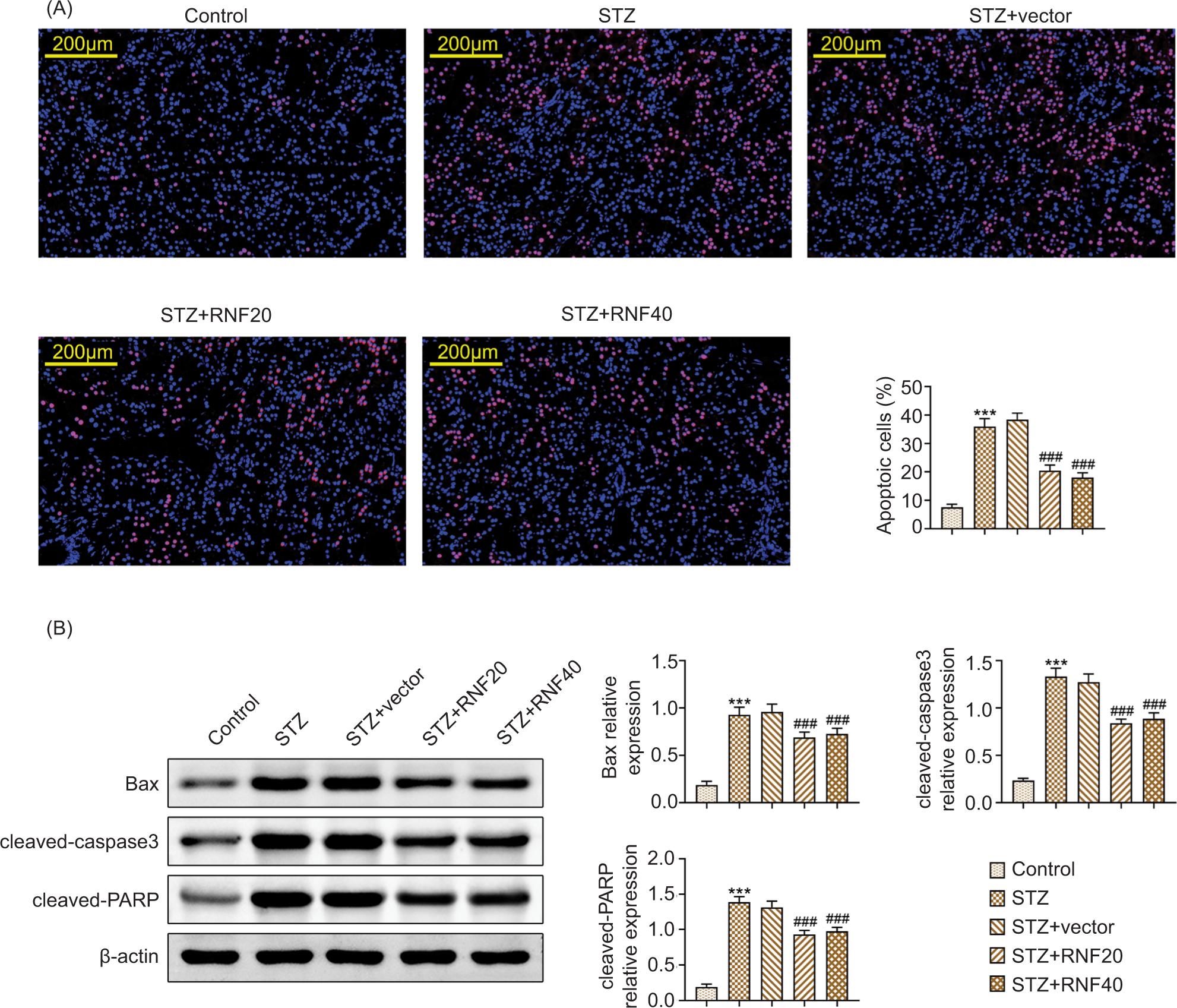
TUNEL assay results revealed that cell apoptosis was promoted in STZ-triggered mice, but this effect was relieved after enhancing RNF20 or RNF40 (Figure 4A). In addition, the increased protein expression of Bax, cleaved caspase-3, and cleaved-PARP mediated by STZ treatment was weakened after RNF20 or RNF40 overexpression (Figure 4B). These findings confirmed that RNF20/RNF40 stimuated cell apoptosis in the pancreatic tissues of STZ-triggered mice.
Figure 4 RNF20/RNF40 improved pancreatic tissue cell apoptosis in STZ-triggered mice. Mice were divided into the following groups: Control, STZ, STZ+vector, STZ+RNF20, and STZ+RNF40. (A) Cell apoptosis in pancreatic tissues was measured through TUNEL assay. (B) The protein expressions of Bax, cleaved caspase-3, and cleaved-PARP were discovered through Western blot analysis. ***P < 0.001 vs the control group; ###P < 0.001 vs the STZ+vector group.
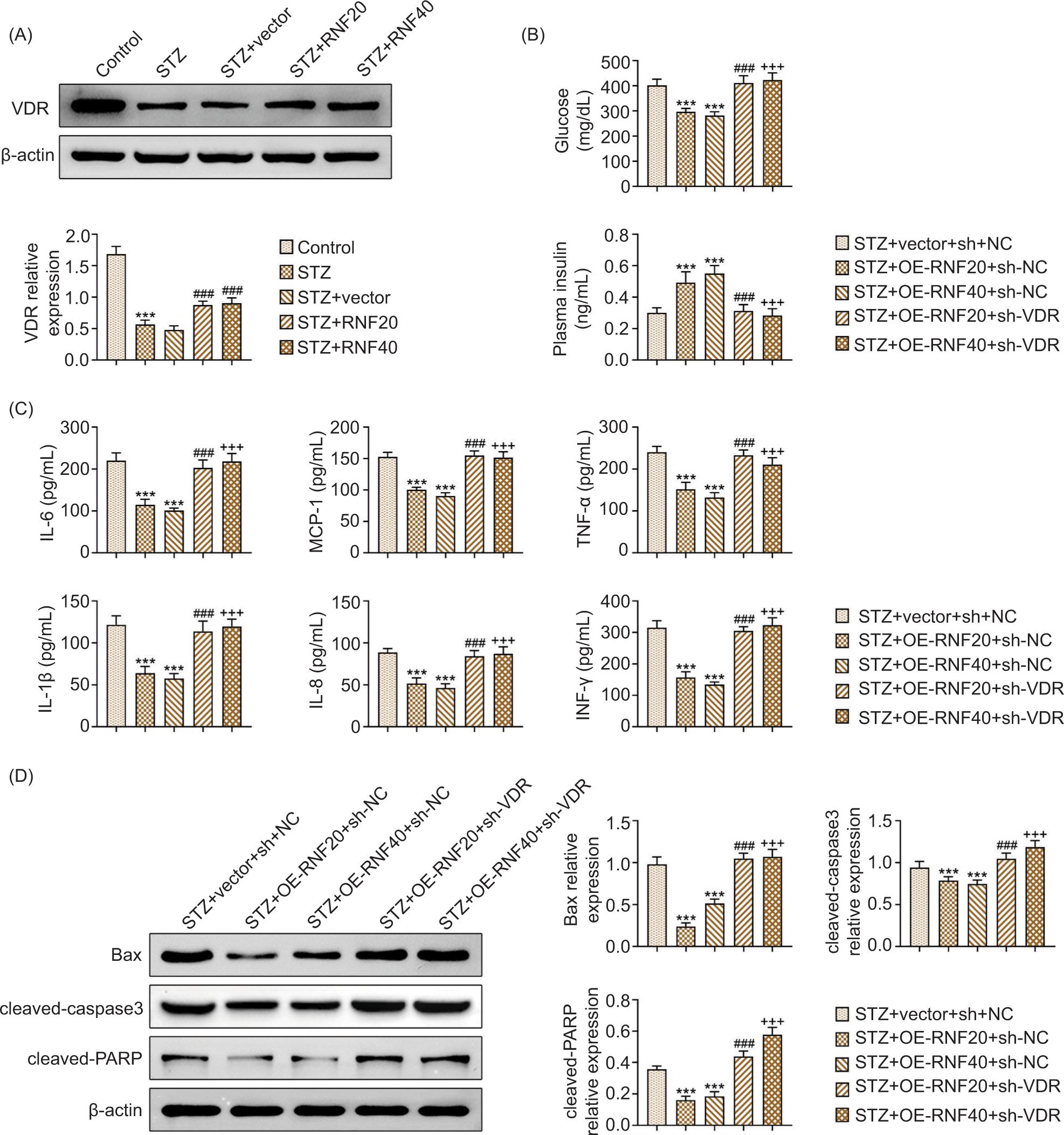
RNF20/RNF40 activated vitamin D receptors to ameliorate type 1 diabetes
In Figure 5A, the decreased protein expression of VDR induced by STZ treatment was saved after enhancing RNF20 or RNF40. Further investigations demonstrated that in the STZ-stimulated mice, glucose decreased and plasma insulin increased after up-regulating RNF20 or RNF40, but these effects were reversed after suppressing VDR (Figure 5B). In the STZ-induced mice, the levels of IL-6, MCP-1, TNF-α, IL-1β, IL-8, and INF-γ were reduced following the overexpression of RNF20 or RNF40; however, these changes were neutralized after inhibiting VDR (Figure 5C). Besides, in the STZ-stimulated mice, the protein expressions of Bax, cleaved caspase-3, and cleaved-PARP were decreased after overexpression of RNF20 or RNF40, but these effects improved after suppressing VDR (Figure 5D). Therefore, RNF20/RNF40 activated vitamin D receptors to relieve type 1 diabetes.
Figure 5 RNF20/RNF40 activated VDRs to ameliorate type 1 diabetes. Mice were divided into the following groups: Control, STZ, STZ+vector, STZ+RNF20, and STZ+RNF40. (A) The protein expression of VDR was examined through Western blot analysis. Groups were divided into the STZ+vector+sh-NC, STZ+RNF20+sh-NC, STZ+RNF40+sh-NC, STZ+RNF20+sh-VDR, and STZ+RNF40+sh-VDR. (B) Fasting blood glucose was measured by glucose meter. The plasma insulin was evaluated using the commercial kit. (C) The levels of IL-6, MCP-1, TNF-α, IL-1β, IL-8, and INF-γ were checked using ELISA. (D) The protein expressions of Bax, cleaved caspase-3, and cleaved-PARP were measured by Western blot analysis. ***P < 0.001 vs the STZ+vector+sh-NC group; ###P < 0.001 vs the STZ+RNF20+sh-NC group; and +++P < 0.001 vs the STZ+RNF40+sh-NC group.
Discussion
RNF20/RNF40 has been examined to take part in various biological processes to result in diseased states. For example, ubiquitin ligase RNF20/RNF40 stabilizes motor protein Eg5 to accelerate spindle assembly, and facilitates breast cancer tumorigenesis.17 Additionally, the RNF20/RNF40 complex affects mRNA splicing and gene transcription by regulating tumor protein p53.18 Besides, RNF20/RNF40 triggers the transcription of heat shock-responsive genes by modulating eEF1BδL monoubiquitylation.19 However, the functioning of RNF20/RNF40 in type 1 diabetes remains unclear. In this work, first, STZ (50 mg/kg) was utilized to stimulate mice model for type 1 diabetes. It was discovered that both RNF20 and RNF40 expressions were down-regulated in STZ-mediated type 1 diabetes. In addition, RNF20/RNF40 improved hyperglycemia in STZ-stimulated mice. Moreover, RNF20/RNF40 lessened pancreatic tissue injury in STZ-induced mice.
Inflammation is a crucial process in the progression of type 1 diabetes. For instance, adipose-derived stem cells suppress mitochondrial stress and inflammation to lessen injury of the liver in type 1 diabetes mellitus.20 Moreover, circular RNA (circRNA) Hsa_circ_0060450 wipes off miR-199-5p to relieve type I interferon-stimulated inflammation in type 1 diabetes mellitus.21 In addition, circRNA circPPM1F accelerates pancreatic islet inflammation and M1 macrophage activation in type 1 diabetes mellitus.22
In the present work, further experiments uncovered that RNF20/RNF40 relieved the strengthened inflammation mediated by STZ treatment. Cell apoptosis was enhanced in the pancreatic tissues of STZ-triggered mice, but this effect was attenuated by the overexpression of RNF20/RNF40.
Vitamin D receptors have been demonstrated as a key regulatory factor in type 1 diabetes. For example, VDR exhibits lower expression in type 2 diabetes, and is closely linked to the severity of albuminuria.23 Furthermore, the H19/miR-675/EGR1 axis down-regulates VDR to participate in the progression of diabetic nephropathy.24
A short of calcium facilitates type 1 diabetes in non-obese diabetic (NOD) mice with lacking VDR.25 Besides, enhancement of VDR in β-cells ameliorates diabetes in mice.26 Importantly, one report indicated that in inflammatory bowel disease, RNF20 and RNF40 can modulate VDR-dependent signaling.16 In this study, the VDR expression was positively regulated by RNF20/RNF40. Finally, it was disclosed that knockdown of VDR reversed the lessened hyperglycemia, inflammation, and cell apoptosis stimulated by the overexpression of RNF20/RNF40.
Studies have shown that loss of RNF20 or RNF40 affects the expression of key genes in pancreatic beta cells. RNF complexes are required for glucose-stimulated insulin secretion and normal mitochondrial ROS levels, suggesting that RNF20 and RNF40 regulate beta cell gene expression and insulin secretion. In addition, it has been shown that RNF20 and RNF40 deletions reduce VDR mRNA and protein levels in inflammatory bowel disease, thereby promoting intestinal inflammation. This study indicated that RNF20/RNF40 activated VDR to ameliorate type 1 diabetes, which further affected apoptosis and inflammation.
Limitation of this study was that although it demonstrated the effect of RNF20/RNF40 on type 1 diabetes progression, the in-depth molecular mechanism studies were lacking. Next, we should use a variety of omics disciplines to screen key signaling pathways and clarify their molecular mechanisms.
Conclusion
In summary, it was the first study to investigate the functioning of RNF20/RNF40 in type 1 diabetes. Our findings disclosed that RNF20/RNF40 activated VDR to ameliorate type 1 diabetes. It established the functioning of RNF20/RNF40 as a promising target for treating type 1 diabetes. In the future, more in-depth studies would be required to explore the regulatory functions of RNF20/RNF40 on other cellular processes in type 1 diabetes.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing Interests
The authors stated that there was no conflict of interest to disclose.
Ethical Approval
Ethical approval was obtained from the Ethics Committee of Ordos Central Hospital (Approval No. 2020-051).
Author Contributions
Junxiang Liu did conceptualization, methodology, and writing of original draft. Xuri Wu and Haixia Qin performed formal analysis, resources, and investigation. Ying Hu carried out formal analysis, visualization, and data curation. Zhiyun Zhang and Yanmei Wang conducted project administration, supervision, and validation. Jinlan Li performed validation, supervision, and writing, including reviewing and editing of paper. All authors read and approved the final manuscript.
REFERENCES
1. Abo El Nazar S, Ghazy AA, Amer I, Tawfik S, Nassar M, Osman EM. Genetic polymorphisms of endoplasmic reticulum amino peptidase 1 (ERAP1) and interferon lambda 4 (IFN-λ4) in Egyptian patients with type 1 diabetes mellitus. Egypt J Immunol. 2023;30:116–24. 10.55133/eji.300112
2. Maahs DM, West NA, Lawrence JM, Mayer-Davis EJ. Epidemiology of type 1 diabetes. Endocrinol Metab Clin North Am. 2010;39(3):481–97. 10.1016/j.ecl.2010.05.011
3. Wright JJ, Williams JM, Letourneau-Freiberg LR, Kandasamy B, Reyes D, Kanegusuku AG, et al. Insulin deficiency from insulin gene mutation leads to smaller pancreas. Diabetes Care. 2023. 10.2337/dc22-2082.
4. Gillespie KM. Type 1 diabetes: Pathogenesis and prevention. Can Med Assoc J (CMAJ). 2006;175(2):165–70. 10.1503/cmaj.060244
5. Li W, Huang E, Gao S. Type 1 diabetes mellitus and cognitive impairments: A systematic review. J Alzheimer’s Dis (JAD). 2017;57(1):29–36. 10.3233/JAD-161250
6. Mahzari M, Al Sohaim A, Al Mulla A, Al Assaf A, Al Amri K, Hussain SD, et al. Variations in cortisol response in patients with known and suspected adrenal insufficiency. Medicine (Baltimore). 2022;101:e32557. 10.1097/MD.0000000000032557
7. Hughes JW, Bao YK, Salam M, Joshi P, Kilpatrick CR, Juneja K, et al. Late-onset T1DM and older age predict risk of additional autoimmune disease. Diabetes Care. 2019;42(1):32–8. 10.2337/dc18-1157
8. Dong JY, Zhang WG, Chen JJ, Zhang ZL, Han SF, Qin LQ. Vitamin D intake and risk of type 1 diabetes: A meta-analysis of observational studies. Nutrients. 2013;5(9):3551–62. 10.3390/nu5093551
9. Sahota O. Understanding vitamin D deficiency. Age Ageing. 2014;43(5):589–91. 10.1093/ageing/afu104
10. Mukhtar M, Batool A, Wajid A, Qayyum I. Vitamin D receptor gene polymorphisms influence T1D susceptibility among Pakistanis. Int J Genomics. 2017;2017:4171254. 10.1155/2017/4171254.
11. Kim J, Guermah M, McGinty RK, Lee JS, Tang Z, Milne TA, et al. RAD6-mediated transcription-coupled H2B ubiquitylation directly stimulates H3K4 methylation in human cells. Cell. 2009;137(3):459–71. 10.1016/j.cell.2009.02.027
12. Shema E, Tirosh I, Aylon Y, Huang J, Ye C, Moskovits N, et al. The histone H2B-specific ubiquitin ligase RNF20/hBRE1 acts as a putative tumor suppressor through selective regulation of gene expression. Genes Dev. 2008;22(19):2664–76. 10.1101/gad.1703008
13. Chernikova SB, Razorenova OV, Higgins JP, Sishc BJ, Nicolau M, Dorth JA, et al. Deficiency in mammalian histone H2B ubiquitin ligase Bre1 (Rnf20/Rnf40) leads to replication stress and chromosomal instability. Cancer Res. 2012;72(8):2111–9. 10.1158/0008-5472.CAN-11-2209
14. Foster ER, Downs JA. Methylation of H3 K4 and K79 is not strictly dependent on H2B K123 ubiquitylation. J Cell Biol. 2009;184(5):631–8. 10.1083/jcb.200812088
15. Wade AK, Liu Y, Bethea MM, Toren E, Tse HM, Hunter CS. LIM-domain transcription complexes interact with ring-finger ubiquitin ligases and thereby impact islet β-cell function. J Biol Chem. 2019;294(31):11728–40. 10.1074/jbc.RA118.006985
16. Kosinsky RL, Zerche M, Kutschat AP, Nair A, Ye Z, Saul D, et al. RNF20 and RNF40 regulate vitamin D receptor-dependent signaling in inflammatory bowel disease. Cell Death Differ. 2021;28(11):3161–75. 10.1038/s41418-021-00808-w
17. Duan Y, Huo D, Gao J, Wu H, Ye Z, Liu Z, et al. Ubiquitin ligase RNF20/40 facilitates spindle assembly and promotes breast carcinogenesis through stabilizing motor protein Eg5. Nat Commun. 2016;7:12648. 10.1038/ncomms13462
18. Wu C, Cui Y, Liu X, Zhang F, Lu LY, Yu X. The RNF20/40 complex regulates p53-dependent gene transcription and mRNA splicing. J Mol Cell Biol. 2020;12(2):113–24. 10.1093/jmcb/mjz045
19. In S, Kim YI, Lee JE, Kim J. RNF20/40-mediated eEF1BδL monoubiquitylation stimulates transcription of heat shock-responsive genes. Nucleic Acids Res. 2019;47(6):2840–55. 10.1093/nar/gkz006
20. Hou Y, Ding W, Wu P, Liu C, Ding L, Liu J, et al. Adipose-derived stem cells alleviate liver injury induced by type 1 diabetes mellitus by inhibiting mitochondrial stress and attenuating inflammation. Stem Cell Res Ther. 2022;13(1):132. 10.1186/s13287-022-02760-z
21. Yang L, Han X, Zhang C, Sun C, Huang S, Xiao W, et al. Hsa_circ_0060450 negatively regulates type I interferon-induced inflammation by serving as miR-199a-5p sponge in type 1 diabetes mellitus. Front Immunol. 2020;11:576903. 10.3389/fimmu.2020.576903
22. Zhang C, Han X, Yang L, Fu J, Sun C, Huang S, et al. Circular RNA circPPM1F modulates M1 macrophage activation and pancreatic islet inflammation in type 1 diabetes mellitus. Theranostics. 2020;10(24):10908–24. 10.7150/thno.48264
23. Yi B, Huang J, Zhang W, Li AM, Yang SK, Sun J, et al. Vitamin D receptor down-regulation is associated with severity of albuminuria in type 2 diabetes patients. J Clin Endocrinol Metab. 2016;101(11):4395–404. 10.1210/jc.2016-1516
24. Fan W, Peng Y, Liang Z, Yang Y, Zhang J. A negative feedback loop of H19/miR-675/EGR1 is involved in diabetic nephropathy by downregulating the expression of the vitamin D receptor. J Cell Physiol. 2019;234(10):17505–13. 10.1002/jcp.28373
25. Driver JP, Lamont DJ, Gysemans C, Mathieu C, Serreze DV. Calcium insufficiency accelerates type 1 diabetes in vitamin D receptor-deficient nonobese diabetic (NOD) mice. Endocrinology. 2011;152(12):4620–9. 10.1210/en.2011-1074
26. Morró M, Vilà L, Franckhauser S, Mallol C, Elias G, Ferré T, et al. Vitamin D receptor overexpression in β-cells ameliorates diabetes in mice. Diabetes. 2020;69(5):927–39. 10.2337/db19-0757