
Download




RESEARCH ARTICLE
Elucidating the probiotic strategy with Pediococcus acidilactici BCB1H and Lactiplantibacillus plantarum HMX2 for restoring normal functions of gut and suppressing inflammation in a DSS-induced colitis animal model
Jasra Naseeba, Muti Ur Rehmanb, Zhennai Yanga*, Tariq Aziza*, Manal F. Elkhadragyc, Wafa A. Alshehrid, Areej A Alhhazmie, Maher S. Alwethaynanif, Hajar Al Qadeebg, Shaza N. Alkhatibh, Khaloud Mohammed Alarjanii, Abeer M. Alghamdij
aKey Laboratory of Geriatric Nutrition and Health of Ministry of Education, Beijing Advanced Innovation Center for Food Nutrition and Human Health, Beijing Technology and Business University, Beijing China
bFood & Biotechnology Research Center (FBRC), Pakistan Council of Scientific Industrial Research (PCSIR), Lahore, Pakistan.
cDepartment of Biology, College of Sciences, Princess Nourah bint Abdulrahman University, PO Box 84428, Riyadh 11671, Saudi Arabia
dDepartment of Food and Nutrition Sciences, College of Agricultural and Food Sciences, King Faisal University, Al Ahsa, Saudi Arabia
eClinical Laboratory Sciences Department, College of Applied Medical Sciences, Taibah University, Medina, Saudi Arabia
fDepartment of Clinical Laboratory Sciences, College of Applied Medical Sciences, Shaqra University, Alquwayiyah, Riyadh, Saudi Arabia
gDepartment of Medical Laboratory, College of Applied Medical Sciences, Prince Sattam Bin Abdulaziz University, Al-Kharj, Saudi Arabia
hDepartment of Biological Sciences, Collage of Sciences and Arts Khulais, University of Jeddah, Jeddah, Saudi Arabia
iDepartment of Botany and Microbiology, College of Science, King Saud University, Riyadh 11451, Saudi Arabia
jDepartment of Biology, Faculty of Science, Al-Baha University, Al-Baha, Saudi Arabia
Abstract
Background: Several inflammatory diseases, including colitis, are treated via probiotics but still the mechanism is not clear.
Objective: This study aimed to restore normal functioning of the gut and ameliorate inflammation in dextran sulfate sodium (DSS)-induced colitis mice via the probiotic intervention of Pediococcus acidilactici (P. acidilactici) BCB1H and Lactiplantibacillus plantarum (L. plantarum) HMX2.
Material and Methods: L. plantarum HMX2 and P. acidilactici BCB1H showed a remarkable reduction in the symptoms of DSS-induced colitis, including weight loss, shortening and damage of the colon, decreased tight junction protein, and pro-inflammatory cytokines in the blood of mice. Two treatment groups were administered with probiotic strains based on animal weight for a duration of 28 days; however, prior to this, DSS was induced.
Results: After consuming both probiotics, the levels of inflammatory markers, such as tumour necrosis factor alpha (TNF-α) and interleukins (IL-1β) exhibited a noticeable decline, indicating a reduction in inflammation. The Disease Activity indices for both probiotic-treated groups (P. acidilactici BCB1H- and L. plantarum HMX2-treated groups) were significantly lower (1±0.45, 1±0.36, 3±0.08, 1±0.18, respectively) than that for the DSS-treated group. Both these treatment strains significantly affected the markers of oxidative stress and inflammation (i.e., malondialdehyde [MDA], superoxide dismutase [SOD], and myeloperoxidase [MPO], compared to the intoxicated group. Similarly, the BCB1H- and HMX2-treated groups showed partial recovery, with improved hepatocyte structure and reduced inflammatory cell infiltration, compared to the intoxicated group. Polymerase chain reaction results of gel analysis showed crucial indicators that measure both colonic integrity and inflammation across four experimental groups of colitis-induced mice. Pro-inflammatory cytokines (TNF-α and IL-1β), E-cadherin, and MUC2 mucin served as evaluated markers across experimental groups.
Conclusion: Increased expression of these genes suggests that both probiotic strains successfully enhanced and restored normal functions of the gut and suppressed inflammation. This could be attributed to the combined immunomodulatory action of probiotics.
Key words: DSS-induced colitis, dysbiosis, gut microbiota, L. plantarum HMX2, P. acidilactici BCB1H
*Corresponding author: Zhennai Yang and Tariq Aziz, Key Laboratory of Geriatric Nutrition and Health of Ministry of Education, Beijing Advanced Innovation Center for Food Nutrition and Human Health, Beijing Technology and Business University, Beijing, China. Email addresses: [email protected], [email protected]
Received 11 August 2025; Accepted 30 September 2025; Available online 1 November 2025
Copyright: Naseeb J, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Probiotics refer to live microorganisms that deliver health advantages after administration of appropriate dosage.1 These microorganisms serve as supplements to improve gut health and performance and adjust the immune system’s functions. The intestinal homeostasis operates through three functions of specific probiotics, such as lactic acid bacteria (LAB), Lactobacillus, Pediococcus acidilactici, and Streptococcus.2 These bacterial species maintain microbiota stability and strengthen mucosal barriers while decreasing inflammation. Scientific research has spotlighted Lactiplantibacillus plantarum (L. plantarum), as it appears in kimchi and yogurt and shows anti-inflammatory properties and generates antioxidants beside anticancer effects. Through its role, intestinal barrier function becomes stronger and microbiota imbalance in inflammatory bowel disease (IBD) shows improvements.3–8 Lactobacillus species have received scientific validation to be safe as probiotics in human subjects and animal models if used to improve intestinal health and the immune system control.9 Laboratory investigations demonstrate that Lactobacillus casei, Lactobacillus paracasei, and Lactobacillus rhamnosus represent the most well-studied probiotics, because these strains exhibit safe behavior while maintaining gut flora health.10,11 The effectiveness of probiotics depends on specific bacterial strains, even though Lplantarum HMX2 and P. acidilactici BCB1H display treatment possibilities for IBD symptoms, but researchers have not established their definite impact on colitis.12 Various studies have demonstrated that L. plantarum differs due to its proven effects on intestinal health in patients with intestinal barrier dysfunction and gut microbiota dysbiosis. It is confirmed from previous studies that intake of L. plantarum delivers significant benefits to patients with intestinal diseases through its dual ability to restore microbial equilibrium and by maintaining intestinal barriers while managing the immune system’s reactions.13 Furthermore, animal studies using dextran sulfate sodium (DSS)-induced colitis models showed that L. plantarum acts as a defense mechanism against gut inflammation and helps to heal the colon.
Different studies have confirmed that the DSS-induced colitis model represents human IBD through its ability to replicate pathophysiological features, such as rise in epithelial permeability breakdown of tight junction (TJ), and elevated cytokine activity.14–18 Studies report that Lactobacillus reuteri (L. reuteri), Lactobacillus fermentum (L. fermentum), and Lactobacillus gasseri (L. gasseri) enhance gut barrier function and influence immune signaling pathways, but scientists have yet to determine their exact mechanisms of action.19,20
Together with L. plantarum, there exists another promising probiotic called P. acidilactici, which demonstrates its ability to strengthen gut barrier function and reduce intestinal inflammatory responses. Two probiotics L. plantarum and P. acidilactici demonstrate their potential to raise TJ protein expressions, including occludin, claudin, and zonula occludens-1, which enhance intestinal barrier strength and inhibit gut permeability. The probiotics generate antioxidant effects to help counter oxidative stress to epithelial cells, thereby promoting the sustainability of homeostasis.21,22 The persistent inflammatory state of IBD affects the gastrointestinal (GI) tract in a continuous and recurring manner. Crohn’s disease (CD), together with ulcerative colitis (UC), forms a major category of IBD, resulting in notable decline in the patient’s quality of life (QoL) through abdominal pain, diarrhea, rectal bleeding, weight loss, and fatigue. The prevalence of IBD has risen globally, particularly in regions where Western diets are common. Factors such as genetic predisposition, environmental influences, dietary habits, gut microbiota imbalance (dysbiosis), and the immune system dysfunction are thought to contribute to disease onset and recurrence.23
The DSS-induced colitis mice model was used to study UC because it develops features of UC biology, including enhanced epithelial permeability with disrupted mucus layer structures, elevated immuno-molecules, and impaired barrier function.24 The pathogenesis of IBD strongly depends on gut microbiota because dysbiosis results in worsened intestinal inflammation and heightened immune system reactions. Dysbiosis causes modifications in epithelial cell functioning and pro-inflammatory cytokine development that boost risks of the disease. The scientific connection between UC and dysbiosis is manifested but the specific nature of this relationship needs further research.25 The initial breakdown of epithelial tissue enables bacteria to enter the body and activates immune responses, leading to severe tissue damage and colitis.26 In addition to microbiota imbalance, oxidative stress is a significant factor in UC. Reduced antioxidants exacerbate intestinal inflammation by increasing pro-inflammatory molecules.27–31 Recent research has highlighted the role of microRNAs (miRNAs), which are associated with the gut microbiota and act as important regulators of intestinal inflammation, immune responses, and mucosal barrier. It is suggested that probiotics could influence miRNA expression, potentially leading to immunomodulatory effects, beneficial in treating IBD.32–35 IBD significantly impacts QoL due to abdominal pain, bloody stools, diarrhea, and weight loss, often leading to emotional distress.36,37 Conventional treatments, such as steroids and amino salicylates, are used commonly, but their long-term use can result in adverse effects. This has led to growing interest in safer and alternative treatments such as probiotics.38,39 This study investigates the anti-inflammatory and probiotic potential of selected L. plantarum HMX2 and P. acidilactici BCB1H strains in a murine DSS-induced colitis model. This research focuses on understanding how L. plantarum HMX2 and P. acidilactici BCB1H modulate genes related to oxidative stress, inflammation, and TJ proteins. Furthermore, the study evaluates how these strains influence the gut microbiome and immune response, particularly by examining the expression of five specific miRNAs associated with colitis.
Material and Methods
Mice grouping, housing, dosing, and establishing DSS-induced colitis
The University of Lahore provided 32 male Albino mice of the same age and weight (75–100 g). Ethical approval for in vivo investigations in this study was granted by PCSIR Laboratories Complex with Ref. No. LLC/PCSIR/24/332. (letter No. 24-332).
The animals were kept in a pathogen-free animal facility having controlled environment, temperature (25±5°C), humidity (60±10%), and a 12-h light and dark cycle with unfettered access to food and water. After that, these animals were bred for 7 days to get used to the new environment. The animals were categorized into four distinct groups i.e. control group (CN), intoxicated negative control DSS-induced colitis (IN), DSS-induced colitis and then treated with P. acidilactici BCB1H (DSS+BCB1H), and DSS-induced colitis and then treated with L. plantarum HMX2 (DSS+HMX2). Other than the CN group, the remaining three groups were subjected to 3.0% DSS induction40,41 (molecular weight: 36–50 kDa; Yeasen Biotechnology Co. Ltd, Shanghai, China) for 8–14 days. Both DSS+BCB1H and DSS+HMX2 groups received 0.2 mL of P. acidilactici BCB1H suspension (1×109–10 CFU/mL) and L. plantarum HMX2 suspension (1×109–10 CFU/mL) via oral gavage once daily from day 11 to day 34. All experimental animal groups were evaluated through weighing them every day along with using a modified Disease Activity Index (DAI) score to track their illness progression. On the morning of day 35, all experimental animals were euthanized.
Physical parameters and disease activity index
Physical parameters involved daily assessments by utilizing a modified DIA scoring system (Table 1), alongside evaluations of stool consistency, weight, and bleeding outcomes in mice participating in DSS experiments. An occult blood detection evaluated presence and absence of blood in examined feces. The colon and other organs were extracted from each animal for histopathological studies. Length of the colon was measured as the first step, followed by proper storage of fecal contents at -20°C.
Table 1 The scoring scale standards.
| Assigned scores | Rectal bleeding | Rectal prolapse | Diarrhea | Colonic bleeding |
|---|---|---|---|---|
| 0 | None | None | No signs | No Signs |
| 1 | Red | Mild | Soft stools | Red |
| 2 | Dark red | Higher | Very soft | Dard red |
| 3 | Gross bleeding | Extensive | Watery stools | Black |
Collection of blood and other samples
After a 28-day experimental study period, mice were euthanized in a fasting state on day 35 at 10 am by administering isoflurane anesthesia. The jugular vein in the neck region of mice was incised using a sharp knife, and blood was collected in sterile tubes for experimental analysis. The blood was collected in tubes containing heparin, which would serve as a material for cytokine and related parameters, and then centrifuged to separate plasma. The plasma was stored in separate portions for examination at a temperature of −20°C.15
Evaluation of MDA, MPO, and SOD
Malondialdehyde (MDA), myeloperoxidase (MPO), and superoxide dismutase (SOD) were evaluated by using locally procured test kits and by following manufacturer’s guidelines (Nanjing Jiancheng Co. Ltd, Nanjing, China). Before being divided into smaller sections, the extracted colonic tissues from each rat were stored in ice-cold buffer at a ratio of 1:10 (w/v). The manufacturer’s specifications guided the tissue homogenization procedure for colonic samples using “IKA” (Staufen, Germany) equipment.
Histopathological study of impairment of colon and liver tissues
Collected colon and liver tissues were preserved after washing with PBS in buffered formalin (neutral, 10%) for histological examinations. Following the procedure outlined by Bancroft and Gamble37, the tissues were divided into thin slices (1 mm) and processed using tissue cassettes in ascending order in graded alcohols to dehydrate the tissue sections. They were then cleared in xylene and stored in paraffin wax to penetrate into tissues and induce support for microtomy. Subsequently, the samples were affixed onto glass slides and subjected to staining using haematoxylin and eosin (H&E) dye. After staining, tissue slides were dehydrated in an ascending order of alcohol (70%, 100% two steps), cleared in xylene (two steps), and mounted with dibutylphthalate polystyrene xylene (DPX). Each slide was covered with glass cover slip (24×50 mm; Marienfeld GmbH, Germany) and observed using a high-power objective lens microscope. Microscopic pictures were acquired for each group at ×40 magnification. Microscopic alterations were assessed and compared with control and negative control groups and those treated with probiotic and symbiotic ice cream samples indicating observable, not observable, and significant damage.
Indicators of inflammation and hepatoprotective parameters
Sterilized tubes were used to collect plasma, which was then evaluated for pro-inflammatory mediators, such as IL-1β and TNF-α, using enzyme-linked-immunosorbent serologic assay (ELISA) kits from MERK (Darmstadt, Germany) using manufacturer’s directions. The data were expressed as pg/mL of total proteins, and the assays were conducted in triplicate.
Gene expression analysis
The process of gene expression analysis, from RNA extraction to reverse transcription PCR (RT-PCR), was conducted following the methodology described by Din et al.43 Briefly, RNA was extracted from tissues and quantified prior to converting into complementary DNA (cDNA) according to the manufacturer’s instructions. Predesigned primers and a PCR kit were used to identify four types of gene expressions with the help of four primers by adhering to the manufacturer’s protocol and as outlined by Din et al.38 PCR reactions were performed using the PCR Master Mix (Thermo Fisher Scientific, USA), incorporating 0.2 μM of each forward and reverse primer along with approximately 10 ng of cDNA template in a 30-μL reaction system. The primers used for the amplification of the target genes, such as IL-1β, TNF-α, MUC-2, and E-cadherin (E-cad), were designed based on sequences retrieved from previous studies and synthesized commercially. Primer sequences, annealing temperatures, and expected amplicon sizes are detailed in Table 2.
Table 2 Primers used in the study.
| Gene | Sequence (5′-3′) | Annealing temperature (ºC) | Product size |
|---|---|---|---|
| IL-1β | FW: CTGAACTCAACTGTGAAATGC RV: TGATGTGCTGCTGCGAGA |
55 | 155 |
| TNF-α | FW: AGGGTCTGGGCCATAGAACT RV: CCACCACGCTCTTCTGTCTAC |
60 | 578 |
| MUC-2 | FW: ATGCCCACCTCCTCAAAGAC RV: GTAGTTTCCGTTGGAACAGTGAA |
56 | 101 |
| E-Cad 1 | FW: CAGGTCTCCTCATGGCTTTGC RV: CTTCCGAAAAGAAGGCTGTCC |
60 | 175 |
Table 3 Comparison of body weight of different treatment groups.
| Body weight in grams/day | Control (n = 8) | Intoxicated group (n = 8) | P. acidilactici BCB1H group(n = 8) | L. plantarumHMX2 group (n = 8) |
|---|---|---|---|---|
| 1 | 75.5±1.18d | 101±1.34a | 88.75±1.26c | 100.83±1.2b |
| 7 | 84.63±1.3d | 105.14±1.02a | 96.87±1.28c | 104.33±1.4b |
| 10 | 93.29±1.42a | 82.57±1.49c | 78.5±1.65d | 84.45±1.58b |
| 15 | 102.7±1.56a | 80±1.37c,d | 81.25±1.12c | 87.53±1.42b |
| 20 | 107.8±1.19a | 78.5±1.28d | 89.25±1.16c | 95.87±1.35b |
| 25 | 113.83±1.22a | 76.52±1.4d | 98.37±1.27c | 101.6±1.3b |
| 30 | 120±1.36a | 77.25±1.31d | 108.25±1.23b | 107.5±1.27b,c |
| 35 | 127.5±1.25a | 76.75±1.06d | 115.5±1.34b | 113.54±1.2c |
Note: Different superscript alphabets within a row represent statistical differences (P < 0.05).
The thermal cycling conditions for PCR were optimized based on specific annealing temperatures of primers. The cycling parameters were as follows:
-
Initial denaturation at 95°C for 5 min
-
Denaturation at 95°C for 30 s
-
Annealing at primer-specific temperature for 30 s
-
Extension at 72°C for 45 s
-
Final extension at 72°C for 7 min
-
Repeated for 30 cycles
The amplified PCR products were mixed with a loading buffer containing SYBR Green dye and subjected to electrophoresis with 2% agarose gel. However, the gel was run at 100 V for 45 min in 1X TAE buffer and subsequently visualized under UV light using a gel documentation system (Bio-Rad, USA). Gene expression levels were determined by the presence and intensity of specific amplicon bands corresponding to expected product sizes. The results were further analyzed for relative expression levels by comparing band intensities using the ImageJ software.
This method allowed for the identification of differential gene expressions in the context of inflammation induced by DSS and its modulation by P. acidilactici BCB1H and L. plantarum HMX2 strains, providing insights into their regulatory effects on inflammatory cytokine profiles.
Statistical analysis
Statistical analysis was conducted using GraphPad Prism ver. 13, with different statistical tests such as one-way ANOVA. The data with P < 0.05 were considered statistically significant.
Results and Discussion
Effect of P. acidilactici BCB1H and L. plantarum HMX2 on physical parameters in DSS-induced colitic mice
The impact of P. acidilactici BCB1H suspension (1×109–10 CFU/mL) and L. plantarum HMX2 suspension (1×109–10 CFU/mL) was assessed on the body weight and DAI of mice model of colitis produced by 3% DSS. As illustrated in Table 3, mice in group CN exhibited robust growth, with a consistent increase in body weight throughout the 35-day trial duration. Body weights of mice in the IN, DSS+BCB1H, and DSS+HMX2 groups were 82.57±1.49 g, 78.5±1.65 g, and 83.45±1.58 g, respectively, on day 10, reflecting a drop from 105.14±1.02 g, 96.87±1.28 g, and 104.33±1.4 g, respectively, before induction on day 7. After the initiation of treatment on day 11 with BCB1H and HMX2 strains, over the next five days (day 15), the CN group gained weight, reaching 102.7±1.56 g, while weight in the intoxicated group dropped further to 80±1.37 g.
Meanwhile, the treated groups showed slight recovery, with P. acidilactici-treated animals having a weight of 81.25±1.69 g and L. plantarum-treated animals having a weight of 87.53±1.8 g. Consequently, on day 20, the body weight increased further, with the CN group attaining a weight of 107.8±1.19 g, while weight in the intoxicated group declined further to 78.5±1.28 g. However, the treated groups exhibited significant recovery, with P. acidilactici recovering a weight of 89.25±1.1 g and L. plantarum recovering a weight of 95.87±1.35 g. This trend continued up to day 25, with the CN group attaining a weight of 113.83±1.22 g, and weight in the intoxicated group dropping to its lowest level of 76.52±1.4 g, and the respective treated groups gaining weight further to 98.37±1.27 g and 101.6±1.3 g. By day 30, the CN group had attained a weight of 120.0 g, and the intoxicated group showing minor improvement at 77.25±1.31 g. The treated groups displayed weight gain, with P. acidilactici at 108.25±1.23 g and L. plantarum at 107.5±1.27 g.
Finally, on day 35, the CN group attained the maximum weight of 127.5±1.25 g while weight in the intoxicated group stagnated at 76.75±1.06 g. Both treatment groups showed substantial recovery, with P. acidilactici-treated group attaining a weight of 115.5±1.34 g and L. plantarum-treated group attaining a weight of 113.54±1.2 g. Overall, the CN group exhibited steady growth, while gradual decrease in the weight was observed in intoxicated (IN) group. The treated groups, with the passage of time, demonstrated effective mitigation of weight gain, which proved that the use of P. acidilactici BCB1H and L. plantarum HMX2 had a preventive function to avert weight loss.44–47
Effects of probiotic treatment on DAI scores after induced DSS
As shown in Table 4, DAI scores of the DSS group for rectal bleeding, rectal collapse, diarrhea, and colonic bleeding were significantly higher (3±0.20, 3±0.12, 3±0.18, 3±0.10, respectively) than that of the CN group on day 8 of DSS intoxication. The symptoms increased up to day 15 and then remained stable relatively. The most common symptoms were rectal prolapse, rectal bleeding, diarrhea, and colonic bleeding.
Table 4 Evaluation of physical parameters and comparison of mean DAI scores in DSS-induced colitis mice.
| Time (day) | Control (n = 8) | Intoxicated group (n = 8) | P. acidilactici BCB1H group (n = 8) | L. plantarum HMX2 group (n = 8) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rectal bleeding, prolapse, diarrhea | Colonic bleeding | Rectal bleeding | Rectal prolapse | Diarrhea | Colonic bleeding | Rectal bleeding | Rectal prolapse | Diarrhea | Colonic bleeding | Rectal bleeding | Rectal prolapse | Diarrhea | Colonic bleeding | |
| 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 8 | 0 | 0 | 1±0.35 | 1±0.15 | 1±0.27 | 1±0.20 | 1±0.31 | 1±0.19 | 1±0.24 | 1±0.16 | 1±0.42 | 1±0.29 | 1±0.34 | 1±0.26 |
| 10 | 0 | 0 | 2±0.51 | 2±0.28 | 2±0.40 | 2±0.64 | 2±0.23 | 2±0.85 | 2±0.54 | 2±0.60 | 2±0.12 | 2±0.25 | 2±0.68 | 2±0.54 |
| 15 | 0 | 0 | 3±0.20 | 3±0.12 | 3±0.18 | 3±0.10 | 1±0.45 | 3±0.08 | 3±0.04 | 3±0.02 | 1±0.36 | 1±0.18 | 1±0.42 | 1±0.52 |
| 20 | 0 | 0 | 2±0.74 | 2±0.56 | 2±0.30 | 2±0.45 | 1±0.16 | 2±0.15 | 2±0.11 | 2±0.18 | 1±0.09 | 1±0.85 | 1±0.30 | 1±0.28 |
| 25 | 0 | 0 | 2±0.42 | 2±0.34 | 2±0.24 | 2±0.18 | 0 | 1±0.12 | 1±0.09 | 1±0.07 | 0 | 1±0.34 | 1±0.11 | 1±0.15 |
| 30 | 0 | 0 | 2±0.68 | 2±0.46 | 2±0.50 | 2±0.54 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 35 | 0 | 0 | 2±0.26 | 2±0.30 | 2±0.35 | 2±0.40 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Effects of probiotics on rectal bleeding and rectal collapse
The DAI index of probiotic-treated groups (P. acidilactici BCB1H and L. plantarum HMX2) was significantly lower (1±0.45, 1±0.36, 3±0.08, 1±0.18, respectively) than that of the DSS-treated group as shown Table 4. In DSS-treated group, we observed increase in rectal bleeding and rectal collapse, which recovered significantly with above-mentioned probiotics.
Effects of probiotics on diarrhea and colonic bleeding
The DAI index scores for diarrhea and colonic bleeding in probiotic-treated groups (P. acidilactici BCB1H and L. plantarum HMX2) were significantly lower (3±0.04, 1±0.42, 3±0.02, 1±0.52, respectively), compared to the DAI index of the DSS-treated group. Diarrhea and colonic bleeding were successfully controlled after treatment with P. acidilactici BCB1H and L. plantarum HMX2. The results showed that probiotic treatment could have reduced the incidence of rectal prolapse, rectal bleeding, diarrhea, and colonic bleeding in intoxicated mice over 35 days of treatment. No symptoms of diarrhea and colonic bleeding were observed at any point in the CN group. In the intoxicated group, symptoms started to appear on day 8 with mild severity (1±0.2–1±0.4) and peaked on day 15 with severe manifestations (3±0.1–3±0.2). Symptoms remained moderate from day 20 to day 35, with values remaining at around 2±0.3–2±0.7.
The P. acidilactici BCB1H-treated group also exhibited mild symptoms starting on day 8, peaking on day 15 (3±0.04–3±0.08 for maximum symptoms) but showed a notable decline by day 20, with rectal bleeding nearly resolved (1±0.16). By day 25, rectal bleeding and colonic bleeding disappeared (value 0), and all symptoms were resolved by day 30. Similarly, the L. plantarum HMX2-treated group followed a comparable trend, with symptoms starting on day 8 and peaking on day 15. However, the symptoms in this group resolved slightly faster, with most symptoms significantly reduced by day 20 (1±0.09–1±0.85) and fully resolving by day 25. Overall, it indicates that both probiotic treatments significantly reduced the severity and duration of symptoms compared to the intoxicated group, with probiotics demonstrating slightly faster resolving of manifestations.48
Evaluation of MDA, MPO, and SOD
The efficiency of a crucial indicator was studied for oxidative stress parameter MDA, inflammatory cells infiltration MPO, and antioxidant enzyme SOD. Probiotic treatment demonstrated powerful competence to minimize inflammatory responses and boost antioxidant/oxidant status by suppressing inflammatory mediators combined with positive effects on catalase (CAT) and SOD antioxidant enzyme activities in colon tissues.49–51
Table 5 shows the effects of different treatments on the markers of oxidative stress and inflammation: MDA, SOD, and MPO. In the CN group, the MDA level was the lowest at 1.06±0.32 nmole/mg, indicating minimal oxidative stress, while SOD activity was moderate at 16.8±2.07 units/g and MPO, a marker of inflammation, was low at 6.28±1.01 units/g, reflecting normal physiological conditions. In contrast, the intoxicated group exhibited significant oxidative stress with a high MDA level of 2.96±0.41 nmole/mg, a sharp reduction in SOD activity to 10.04±1.81 units/g, and an elevated MPO level of 14.08±1.63 units/g, indicating increased inflammation and compromised antioxidant defenses. Treatment with P. acidilactici BCB1H resulted in a moderate reduction of oxidative stress, as evidenced by an MDA level of 1.575±0.30 nmole/mg, and a significant enhancement of antioxidant activity with an SOD level of 23.875±5.52 units/g, alongside a reduction in inflammation, as reflected by an MPO level of 7.4625±1.24 units/g. Similarly, treatment with L. plantarum HMX2 led to an MDA level of 2.35±0.94 nmole/mg, indicating partial protection against oxidative stress, while achieving the highest SOD activity at 26.3±3.45 units/g and a relatively low MPO level of 6.95±0.38 units/g, suggesting reduced inflammation.
Table 5 Effects of probiotic treatment on MDA, MPO, and SOD levels in DSS-induced colitis mice.
| Group | MDA (nmole/mg) | SOD (units/g) | MPO (units/g) |
|---|---|---|---|
| Control (n = 8) | 1.06±0.32d | 16.8±2.07c | 6.28±1.01c,d |
| Intoxicated group (n = 8) | 2.96±0.41a | 10.04±1.81d | 14.08±1.63a |
| P. acidilactici BCB1H group (n = 8) | 1.575±0.30c | 23.875±5.52b | 7.4625±1.24b |
| L. plantarum HMX2 group | 2.35±0.94b | 26.3±3.45a | 6.95±0.38b,c |
Note: Different superscript alphabets within a column represent statistical differences (P < 0.05).
Overall, while both treatments demonstrated protective effects , P. acidilactici BCB1H was more effective in reducing oxidative stress and inflammation, whereas L. plantarum HMX2 provided the strongest boost to antioxidant defenses.
Effects on inflammatory cytokines
Figure 1 shows that mice in the intoxicated group produced higher TNF-α and IL-1β levels, marking increased inflammatory activity. The cytokines signaling pathway controls inflammatory signal transduction pathways that help to reduce UC. These cytokines lead to intestinal inflammation through the release of inflammatory mediators.52 Management of intestinal homeostasis depends on proper pro-inflammatory cytokines regulation throughout the colonic mucosal tissue. The production of abnormal pro-inflammatory cytokines disrupts balance in patients with IBD, specifically CD and UC.53 Findings of the current study showed meaningful evidence of inflammatory response detection through gene expression examination of the colon. Laboratory investigators determined the levels of TNF and IL-1β present in blood serum. Pro-inflammatory cytokines have been shown to cause damage to epithelial TJs, leading to increased intestinal permeability.54,55 The BCB1H- and HMX2-treated groups had lower levels of TNF-α and IL-1β, indicating reduced inflammation that helped to improve colitis-induced permeability disruption by suppressing inflammatory cytokine production (Figure 1). Active signaling pathways are triggered by toll-like receptors (TLRs) that control the expression of inflammation and immune response-related genes.56
Figure 1 Level of cytokines in different treatment groups. Different superscript alphabets in a column represent statistical differences (P < 0.05).
The general microbial diversity found in IBD patients remains lower than that exhibited in healthy people.57 The research findings demonstrated that DSS treatment resulted in α-diversity reduction, which is measured by Chao1, observed and ACE indicators i.e. in the gut (colon), the Chao1 and ACE are observed indicators used to measure α-diversity (alpha-diversity), which is a measure of the total number of species and the overall diversity of the microbial community. Although probiotic treatment did not restore α-diversity measurements to normal, it managed to normalize α-diversity into ranges equivalent to the CN group. Examination of β-diversity revealed distinct groups between control and DSS-treated subjects, yet the probiotic-treated cohort had compositional similarities to the CN group, rather than the DSS group. The pathogenic bacterial genera within Proteobacteria, such as Escherichia, Enterobacter, Vibrio, Yersinia, and Salmonella, disrupt intestinal microbial composition in both colitis models and IBD patients.58 Among these, Escherichia (part of Proteobacteria phylum and Enterobacteriaceae family) are prominent pathogens that exacerbate intestinal inflammation and contribute to bacterial dysbiosis in the mucosa of mice with colitis.58,59 The BCB1H- and HMX2-treated groups had lower levels of TNF-α and IL-1β, indicating reduced inflammation. Probiotic treatments (BCB1H and HMX2) have anti-inflammatory effects, mitigating the increased inflammation caused by toxic substances.
Microscopic and gross histopathological comparison of colon
The histological examination of mice colon across all experimental groups yielded the following findings. Colon segments in the CN group (Figure 2A) displayed a histological structure that conformed to normal values. This encompassed the mucosa (composed of Lamina muscularis mucosa, Lamina epithelialis, and lamina propria), submucosa, muscular coat (consisting of inner circular and outer longitudinal muscle fibers), and serosa. In the DSS-induced colitis group (Figure 2B), colon segment exhibited numerous ulcerative regions in mucosal layers, accompanied by considerable tissue damage. Furthermore, a significant inflammatory reaction accompanied by congestion was noted in the blood vessels of both mucosal and submucosal layers. The investigation disclosed significant cellular necrosis and desquamation of both superficial and deep epithelial cells, alongside the infiltration of leukocytes, specifically lymphocytes and a limited quantity of neutrophils. Intestinal barrier dysfunction occurred when increased permeability compromised TJ barrier.60,61 Colonic glands displayed localized abnormalities and indicated an absence of secretory function. Colon length was a representative marker for evaluating colitis-induced mice models.62
Figure 2 Effects of probiotics on the histological structure of the colon and the mucosa of the intestine: (A) control; (B) intoxicated group, and (C and D) treatment groups after DSS-induced colitis. Histological sections were stained with H&E (magnification 40×).
The treatment groups receiving two different probiotic combinations following disease induction (Figures 2C and 2D) presented with regular histological features of mucosal and submucosal as well as muscular and serosal layers of their colon sections (Figures 2C and 2D). Studies revealed that every examined group presented recuperated ulcerative lesions together with mucosal epithelium regeneration. Focal hyperplastic changes alongside hyperchromatic nuclei occurred in the newly formed epithelial cells because of regenerative signals. Area near the healed ulcer presented heavy inflammation because plasma cells and lymphocytes accumulated due to its proximity to the mucosa. A large amount of aggregated and/or follicular-type lymphoid cells extensively invaded the submucosa.
The data in Table 6 show the mean colon length (in inches) for various experimental groups, highlighting the effects of treatments on colon health.
Table 6 Mean colon length (in inches) for various experimental groups, highlighting the effects of treatments on colon health.
| Group name | Colon length (inches) |
|---|---|
| Control (CN) | 7.6±0.45 (40%) |
| Intoxicated (IN) | 5.5±0.3 (10%) |
| BCB1H-treated group | 6.7±0.34 (20%) |
| HMX2-treated group | 6.9±0.42 (30%) |
The CN group had the longest mean colon length, measuring 7.6±0.25 inches, representing normal, healthy conditions. In comparison, the intoxicated group displayed a significantly shorter mean colon length of 5.5±0.3 inches, indicating substantial colon damage or shrinkage due to intoxication. Treatment with P. acidilactici BCB1H improved the mean colon length to 6.7±0.34 inches, demonstrating considerable recovery, although not to the level of the CN group. Similarly, treatment with L. plantarum HMX2 resulted in a mean colon length of 6.9±0.42 inches, reflecting slightly better recovery, compared to the BCB1H treatment. Overall, both treatments showed protective effects against intoxication-induced colon shortening, with L. plantarum HMX2 offering slightly greater restoration than P. acidilactici BCB1H. Figures 2C and 2D show the effects of probiotics on colon length across various experimental groups after DSS-induced colitis, with a ruler used for measurement. In the CN group, the colon appeared to be of normal length, indicating a healthy, unaffected state. In contrast, the intoxicated group showed noticeable colon shortening and irregularities, which were characteristic of DSS-induced colitis because of inflammation and tissue damage. The P. acidilactici BCB1H-treated group exhibited partial restoration of colon length, reflecting some recovery, compared to the intoxicated group. The L. plantarum HMX2-treated group showed an even greater improvement, with the colon length nearly restored to normal, suggesting a stronger protective and reparative effect. Overall, Figure 5 highlights the beneficial impact of probiotics, particularly HMX2, in reducing colitis-related damage and promoting colon recovery. Figures 2A–D demonstrate histopathological alterations in colonic and intestinal mucosa structure inside experimental DSS-induced colitis groups that received probiotics therapy. H&E stain was used for sections, which were examined under 40× magnification. Samples from Control group presented normally structured colon tissue alongside intact epithelial layers and organized crypts. The tissue surface displayed regular smooth surfaces without indications of inflammatory damage. The severe histological damage found in intoxicated group included colon tissues exposed to DSS following alcohol intoxication, destruction of epithelial layers combined with irregular crypt structure along with visible tissue inflammation through mucosal damage and invasive inflammatory cells.
Figure 3 Different levels of liver damage reflecting treatment outcomes in experimental groups through histopathological tissue examinations.
Figure 4 RT-qPCR analysis of miRNA expression profiles in mice subjected to DSS-induced colitis and treated with probiotic strains. The relative fold expressions of (A) miR-143, (B) miR-150, (C) miR-223, (D) miR-155, and (E) miR-375 were determined across four experimental groups of control (CN), DSS-induced colitis (IN), DSS+P acidilactici BCB1H (BCB1H), and DSS+L. plantarum HMX2 (HMX2). The data were subjected to statistical analysis, and miRNA levels were normalized to endogenous controls and expressed as mean±SD (n = 8/group). The data were subjected to analysis of variance (ANOVA) to determine statistical significance. The degree of significance was indicated by notations *P = 0.05, **P = 0.001, and ***P = 0.0001, while the absence of any notation indicated lack of statistical significance.
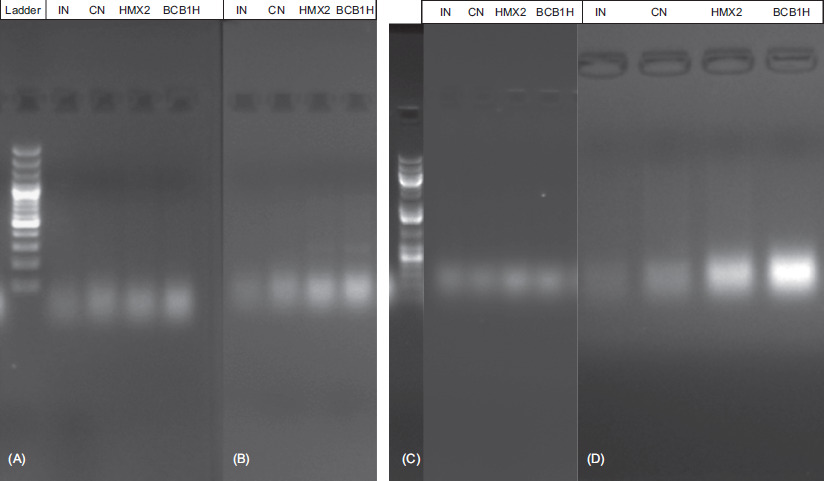
Figure 5 Gel electrophoresis through PCR amplifications to assess the gene expression of two markers (A) TNF-α and (B) E-cadherin (E-cad), and the genes (C) MUC2 and (D) IL-1β.
Histological structural recovery was substantial in animal taking probiotic treatment (BCB1H-treated group and HMX2-treated group), compared to the untreated intoxicated group. The BCB1H-treated group demonstrated partial epithelial repair along with structured crypt organization, whereas group D showed nearly full restoration of epithelial tissue together with normal crypt morphology. Both groups demonstrated decreased inflammation, but HMX2-treated group presented the most noticeable improvement together with better tissue structural normalization. The probiotic-treated groups (BCB1H-treated group and HMX2-treated group) exhibited reduced infiltration of inflammatory cells, compared to the intoxicated group, with fewer neutrophils and a more balanced presence of macrophages and lymphocytes, suggesting healing and resolution of inflammation. IBD reduces the production of short-chain fatty acids, such as butyrate. Butyrate-producing bacteria are associated with increased intestinal homeostasis and reduced inflammation.63–65
Microscopic and gross histopathological comparison of the liver
Figure 3 depicts histological sections of the liver stained with H&E from four experimental groups, highlighting the anti-inflammatory effects of probiotics after DSS-induced damage. In the CN group, the liver showed a normal histological structure with intact hepatocytes and well-organized sinusoids, indicating healthy, undamaged tissue. Conversely, the intoxicated group displayed significant liver damage, including disrupted hepatocyte architecture, cellular swelling, and marked infiltration of inflammatory cells, such as neutrophils and macrophages, indicative of an active inflammatory response caused by DSS exposure. The P. acidilactici BCB1H-treated group showed partial recovery, with improved hepatocyte structure and reduced inflammatory cell infiltration compared to the intoxicated group. This suggests a slight restoration of liver integrity and moderate anti-inflammatory effect because of probiotic intervention. The HMX2-treated group showed the most substantial improvement, with hepatocyte organization largely restored and minimal inflammatory cell presence, closely resembling the CN group’s healthy tissue. These findings concluded that L. plantarum HMX2 exerts a stronger anti-inflammatory and hepatoprotective effect compared to P. acidilactici BCB1H. The liver histology illustrated a potential for probiotics, particularly HMX2, in lessening inflammation and promoting recovery of tissue from DSS-induced liver damage.
Implications of gene expression by RT-qPCR analysis of treated groups and comparison of gel electrophoresis
Quantitative real-time qPCR analysis of miRNA expression profiles in mice subjected to DSS-induced colitis and treated with probiotics are presented in Figures 4A–E. The objective was to assess the modulation of key colonic miRNAs involved in inflammation and immune regulation because of probiotic interventions. The expression of miR-143 (Figure 4A) was markedly suppressed in the DSS-induced IN group, compared to the CN group, reflecting its potential role in preserving mucosal integrity under physiological conditions. Restoration of its levels following both probiotic treatments, particularly with L. plantarum HMX2, suggested a beneficial anti-inflammatory effect and potential mucosal healing activity. miR-150 (Figure 4B), which plays a pivotal role in immune regulation, showed a notable increase in the DSS-induced (IN) group, indicative of immune system dysregulation induced by DSS. Administration of both P. acidilactici and L. plantarum significantly reduced miR-150 expression, demonstrating the probiotics’ capacity to attenuate the heightened immune response associated with colitis. miR-223 (Figure 4C) was significantly upregulated in DSS-treated mice (IN), in alignment with its known function in promoting neutrophil activation and inflammation. Interestingly, treatment with P. acidilactici (BCB1H) led to a more substantial downregulation of miR-223 than that with L. plantarum, highlighting a potentially stronger immunomodulatory action of BCB1H.
Similarly, miR-155 (Figure 4D), a well-established pro-inflammatory miRNA, exhibited elevated expression in the DSS-induced (IN) group, reinforcing its central role in colitis pathology. Both probiotic strains were able to mitigate this increase, with a particularly marked reduction observed in the BCB1H group, supporting its role in dampening inflammatory pathways. In contrast, miR-375 (Figure 4E) displayed minimal fluctuation across all groups, indicating that this miRNA may not be directly involved in DSS-induced inflammation or that its expression is not responsive to probiotic intervention. These findings reinforced the utility of the DSS-induced colitis model in simulating IBD by producing dysregulated miRNA expression profiles that mirror clinical observations. Moreover, the observed restoration of miRNA balance through probiotic administration—especially with P. acidilactici—provided compelling evidence of their therapeutic potential. Altogether, these miRNAs not only contribute to the understanding of molecular mechanisms underlying colitis but also emerge as promising biomarkers and therapeutic targets for IBD intervention strategies. The gel electrophoresis analysis in Figures 5 presents examination results of crucial indicators that measure both colonic integrity and inflammation across four experimental groups of DSS-induced colitis mice. Pro-inflammatory cytokines (TNF-α and IL-1β), E-cad, and MUC2 mucin served as evaluated markers across all experimental groups.
Results in Figure 5A indicate an increased expression of MUC2 in the groups treated with P. acidilactici BCB1H and L. plantarum HMX2, compared to the intoxicated negative control (IN). This increased expression suggests that both probiotic strains successfully enhanced the production of colonic mucin, a critical component of mucus barrier. The restoration of MUC2 levels is indicative of the strains’ potential in mitigating the deterioration of mucus barrier typically observed in UC. These probiotics may indirectly reduce susceptibility to inflammation by fortifying mucus barrier and by enhancing protection against luminal antigens and pathogens. The RT-qPCR analysis revealed a significant downregulation of IL-1β and IL-6 transcription in probiotic-treated groups (L. plantarum HMX2 and P. acidilactici BCB1H), thus aligning with their proposed anti-inflammatory roles. However, the expression of TNF-α did not show significant changes across the groups. This subtle effect on TNF-α might suggest strain-specific or localized regulation of this cytokine, highlighting the complex interplay between probiotics and inflammatory mediators. Similarly, E-cad, a key marker of epithelial integrity, was significantly downregulated in the intoxicated negative control group, indicating epithelial damage because of DSS-induced colitis. However, its expression levels were restored in the probiotic-treated groups, suggesting the potential role of P. acidilactici and L. plantarum in reinforcing epithelial cell–cell adhesion and reducing epithelial barrier dysfunction. The results underlined the potential of both P. acidilactici BCB1H and L. plantarum HMX2 in repairing colonic damage in DSS-induced colitis. Their administration not only upregulated protective markers such as MUC2 and E-cad but also downregulated key pro-inflammatory cytokines, with the exception of TNF-α. The enhancement of mucus barrier and epithelial integrity probably contributes to their anti-inflammatory effects, making these strains promising candidates for therapeutic intervention in UC.
Figures 5A,B display gel electrophoresis results from PCR amplifications to assess the gene expression of two markers: TNF-α and E-cad. Panel A shows the expression of the TNF-α gene, which is associated with inflammation. The lanes represent different experimental groups: IN (intoxicated negative control), CN (control), HMX2 (treated with L. plantarum HMX2), and BCB1H (treated with P. acidilactici BCB1H). The presence of bands indicates the expression of TNF-α, with the intensity of the bands potentially correlating with the level of cytokine production. Panel B examines the expression of E-cad, a protein crucial for maintaining epithelial integrity. The bands in this panel reflect how both probiotics (HMX2 and BCB1H) could influence E-cad expression, which would help to restore epithelial barrier function in DSS-induced colitis. The 1-kb plus DNA ladder on the left serves as a reference for determining the size of PCR products.
Gel electrophoresis results from PCR amplification of the MUC2 and IL-1β genes are depicted in components (Figures 5C and 5D). It evaluates gene expression related to DSS-induced colitis and probiotic treatments. The MUC2 gene expression data can be viewed in image C, as it is vital for upholding the protective intestinal mucus barrier. Different experimental groups tested within Figures 5C and 5D include intoxicated negative control, control, HMX2, and BCB1H. The presence of bands demonstrates MUC2 production, thus showing how probiotics affect operations of intestinal barrier. Band intensities indicate the amount of genetic material produced by the cells. The gel image (Figure 5D) showed the amplified levels of IL-1β, an inflammatory cytokine. The bands in this panel demonstrate the levels of the IL-1β gene expression to determine gut inflammatory responses. The IL-1β gene expression patterns increased with high measures of inflammation but decreased with probiotic-mediated anti-inflammatory effects. By analyzing DNA products through the reference 100 bp plus DNA ladder, scientists are able to determine their sizes. The study proved the understanding of how probiotics affect MUC2 expression while controlling inflammatory responses through IL-1β to demonstrate their potential therapeutic effects on gut inflammation and intestinal health support.
Conclusion
The human immune system and its immunological responses are significantly influenced by gut microbiota, a complex community of billions of bacteria residing in the GI tract. Gut microbiota plays a crucial role in defending against pathogens and is closely associated with various inflammatory diseases, including IBD. Current treatments of IBD are often ineffective or come with undesirable adverse effects; hence, no definitive cure is available. Therefore, the search for a more effective therapeutic approach remains critical. While probiotics have shown promise in managing IBD, their effects on composition of gut microbiome and immunomodulatory mechanisms in murine colitis models are underexplored. This study investigated, for the first time, the therapeutic potential of selected L. plantarum HMX2 and P. acidilactici BCB1H strains and evaluated their impact on gut microbiome dysbiosis, inflammatory cytokines, oxidative stress, intestinal integrity, and liver histopathology, implicating that both these probiotics helped to restore normal state in mice.
Ethical Approval
Ethical approval for this study was granted by PCSIR Laboratories Complex under Ref. No. LLC/PCSIR/24/332.
Competing Interests
The authors declared no conflict of interest.
Availability of Data and Materials
All the data generated in this research work are included in the manuscript.
Author Contributions
Conceptualization: Zhennai Yang and Tariq Aziz; methodology: Jasra Naseeb; software: Abeer M. Alghamdi; validation: Manal F. Elkhadragy and Fahad Al-Asmari; formal analysis: Khaloud Mohammed Alarjani; investigation: Jasra Naseeb and Muti Ur Rehman; resources: Zhennai Yang; data curation: Shaza N. Alkhatib; writing—original draft preparation: Maher S. Alwethayanani; writing—review and editing: Hajar Al Qadeeb; visualization: Areej A Alhhazmi; supervision: Zhennai Yang and Tariq Aziz; and project administration: Zhennai Yang.
Funding
This research was financially supported by the National Natural Science Foundation of China (Project No. 32272296). The authors are thankful to Princess Nourah bint Abdulrahman University Riyadh, Saudi Arabia and its Researchers Supporting Project No. PNURSP2025R23. The authors expressed their gratitude to the Deanship of Scientific Research (DSR) at King Faisal University under Project No. KFU250665.
REFERENCES
1 Sanders ME, Gibson, G., Gill HS, Guarner F. Probiotics: Their potential to impact human health. Counc Agric Sci Technol Issue Pap. 2007;36:1–20.
2 Amelia R, Philip K, Pratama YE, Purwati E. Characterization and probiotic potential of lactic acid bacteria isolated from dadiah sampled in West Sumatra. Food Sci Technol. 2020;41:746–52. 10.1590/fst.30020
3 Kim KJ, Paik H-D, Kim JY. Immune-enhancing effects of Lactiplantibacillus plantarum 200655 isolated from Korean kimchi in a cyclophosphamide-induced immunocompromised mouse model. J Microbiol Biotechnol 2021;31:726–32. 10.4014/jmb.2103.03028
4 Zhou X, Zhang D, Qi W, Hong T, Xiong T, Wu T, et al. Exopolysaccharides from Lactiplantibacillus plantarum NCU116 facilitate intestinal homeostasis by modulating intestinal epithelial regeneration and microbiota. J Agric Food Chem. 2021;69:7863–73. 10.1021/acs.jafc.1c01898
5 Duary RK, Bhausaheb MA, Batish VK, Grover S. Anti-inflammatory and immunomodulatory efficacy of indigenous probiotic Lactiplantibacillus plantarum Lp91 in colitis mouse model. Mol Biol Rep. 2012;39:4765–75. 10.1007/s11033-011-1269-1
6 Ma Y, Fei Y, Han X, Liu G, Fang J. Lactiplantibacillus plantarum alleviates obesity by altering the composition of the gut microbiota in high-fat diet-fed mice. Front Nutr. 2022;9:947367. 10.3389/fnut.2022.947367
7 Li C, Nie S-P, Zhu K-X, Ding Q, Li C, Xiong T, et al. Lactiplantibacillus plantarum NCU116 improves liver function, oxidative stress and lipid metabolism in rats with high fat diet induced non-alcoholic fatty liver disease. Food Funct. 2014;5:3216–23. 10.1039/C4FO00549J
8 Jeong S, Kim Y, Park S, Lee D, Lee J, Hlaing SP, et al. Lactiplantibacillus plantarum metabolites elicit anticancer effects by inhibiting autophagy-related responses. Molecules. 2023;28:1890. 10.3390/molecules28041890
9 Zhang C-X, Wang H-Y, Chen T-X. Interactions between intestinal microflora/probiotics and the immune system. Biomed Res. Int. 2019;2019:919. 10.1155/2019/6764919
10 Jang YJ, Kim W-K., Han, D. H., Lee, K., Ko, G. Lactobacillus fermentum species ameliorate dextran sulfate sodium-induced colitis by regulating the immune response and altering gut microbiota. Gut Microbes. 2019;10:696–711. 10.1080/19490976.2019.1589281
11 Hill D, Sugrue I, Tobin C, Hill C, Stanton C., Ross RP. The Lactobacillus casei group: History and health related applications. Front Microbiol. 2018;9:107. 10.3389/fmicb.2018.02107
12 Gill HS, Guarner F. Probiotics and human health: A clinical perspective. Postgrad Med J. 2004;80:516–26. 10.1136/pgmj.2003.008664
13 Wang J, Ji H, Wang S, Liu H, Zhang W, Zhang D, et al. Probiotic lactiplantibacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Front Microbiol. 2018;9:1953. 10.3389/fmicb.2018.01953
14 Ewaschuk JB, Dieleman LA. Probiotics and prebiotics in chronic inflammatory bowel diseases. World J Gastroenterol WJ. 2006;12:5941. 10.3748/wjg.v12.i37.5941
15 Li Q, Zheng T, Ding H, Chen J, Li B, Zhang Q, et al. Exploring the benefits of probiotics in gut inflammation and diarrhea—from an antioxidant perspective. Antioxidants. 2023;12:1342. 10.3390/antiox12071342
16 Chassaing B, Aitken JD, Malleshappa M, Vijay-Kumar M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr Protoc Immunol. 2014;104:15–25. 10.1002/0471142735.im1525s104
17 Eichele DD, Kharbanda KK. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J Gastroenterol. 2017;23:6016. 10.3748/wjg.v23.i33.6016
18 Kanwal S, Joseph TP, Aliya S, Song S, Saleem MZ, Nisar MA, et al. Attenuation of DSS-induced colitis by dictyophoraindusiata polysaccharide (DIP) via modulation of gut microbiota and inflammatory-related signaling pathways. J Funct Foods. 2020;64:103641. 10.1016/j.jff.2019.103641
19 Zhang W-Q, Quan K-Y, Feng C-J, Zhang T, He Q-W, Kwok L-Y, et al. The Lactobacillus gasseri G098 strain mitigates symptoms of DSS-induced inflammatory bowel disease in mice. Nutrients. 2022;14:3745. 10.3390/nu14183745
20 Sun M-C, Zhang F-C, Yin X, Cheng B-J, Zhao C-H, Wang Y-L, et al. Lactobacillus reuteri F-9-35 prevents DSS-Induced colitis by inhibiting proinflammatory gene expression and restoring the gutmicrobiota in mice. J Food Sci. 2018;83:2645–52. 10.1111/1750-3841.14326
21 Guo N, Lv L. Mechanistic insights into the role of probiotics in modulating immune cells in ulcerative colitis. Immun Inflamm Dis. 2023;11:e1045. 10.1002/iid3.1045
22 Kim H, Yoo M-S, Jeon H, Shim J-J, Park W-J, Kim J-Y, et al. Probiotic properties and safety evaluation of Lactiplantibacillus plantarum HY7718 with superior storage stability isolated from fermented squid. Microorganisms. 2023;11:2254. 10.3390/microorganisms11092254
23 Molodecky NA, Soon IS, Rabi DM, Ghali WA, Ferris M, Chernoff G, et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology. 2012;142:46–54.e42. 10.1053/j.gastro.2011.10.001
24 Alemany-Cosme E, Sáez-González E, Moret I, Mateos B, Iborra M, Nos P, et al. Oxidative stress in the pathogenesis of Crohn’s disease and the interconnection with immunological response, microbiota, external environmental factors, and epigenetics. Antioxidants (Basel). 2021;10:64. 10.3390/antiox10010064
25 Haneishi Y, Furuya Y, Hasegawa M, Picarelli A, Rossi M., Miyamoto J. Inflammatory bowel diseases and gut microbiota. Int J Mol Sci. 2023;24:817. 10.3390/ijms24043817
26 Buttó LF, Haller D. Dysbiosis in intestinal inflammation: Cause or consequence. Int J Med Microbiol. 2016;306:302–9. 10.1016/j.ijmm.2016.02.010
27 Guenther C, Josenhans C, Wehkamp J. Crosstalk between microbiota, pathogens and the innate immune responses. Int J Med Microbiol. 2016;306:257–65. 10.1016/j.ijmm.2016.03.003
28 Chen D, Lv J, Han T, Kan J, Jin CH, Liu J. Hepatic antioxidant and gut ecological modulation properties of long-term intake of tea (Camellia sinensis L.) flower extract in vivo. Quality Assurance and Safety of Crops & Foods. 2023;15(3):11–21. 10.15586/qas.v15i3.1209
29 Song W, Lv W, Bi N, Wang G. Tectorigenin suppresses the viability of gastric cancer cells in vivo and in vitro. Quality Assurance and Safety of Crops & Foods. 2023;15(3):117–125. 10.15586/qas.v15i3.1357
30 Aziz T, Shabbir MA, Sarwar A, Yang Z, Lin L, Al-Megrin WAI, Shami A, Alwethaynani MS, Alhhazmi AA, Al-Asmari F, Al-Joufi FA, Fallatah D. Revealing Lactiplantibacillus Plantarum K25 Derived (Z)-18-Octadec-9-Enolide in Modulating Aquaporin-8 in Colorectal Cancer by Bridging the Gut Microbiome and Membrane Biology. Cell Biochem Biophys. 2025;1–16. 10.1007/s12013-025-01853-9
31 Vargas Robles H, Citalán Madrid AF, García Ponce A, Silva Olivares A, Shibayama M, Betanzos A, et al. Experimental colitis is attenuated by cardioprotective diet supplementation that reduces oxidative stress, inflammation, and mucosal damage. Oxidative Med Cell Longev. 2016;2016:1–9. 10.1155/2016/8473242
32 Wang C, Chen J. microRNAs as therapeutic targets in intestinal diseases. ExRNA 2019;1:1–12. 10.1186/s41544-019-0026-9
33 Ul Ain N, Naveed M, Aziz T, Shabbir MA, Al Asmari F, Abdi G, Sameeh MY, Alhhazmi AA. Mix-match synthesis of nanosynbiotics from probiotics and prebiotics to counter gut dysbiosis via AI integrated formulation profiling. Sci Rep. 2024;8;14(1):18397. 10.1038/s41598-024-69515-z
34 Ahmad W, Din AU, Khan TM, Rehman MU, Hassan A, Aziz T, Alharbi M, Wu J. Lacticaseibacillusparacasei BNCC345679 revolutionizes DSS-induced colitis and modulates gut microbiota. Front Microbiol. 2024;15:1343891. 10.3389/fmicb.2024.1343891
35 Algieri F, Garrido-Mesa J, Vezza T, Rodríguez-Sojo MJ, Rodríguez-Cabezas ME, Olivares M, et al. Intestinal anti-inflammatory effects of probiotics in DNBS-colitis via modulation of gut microbiota and microRNAs. Eur J Nutr. 2021;60:2537–51. 10.1007/s00394-020-02441-8
36 Seyedian SS, Nokhostin F, Malamir MD. A review of the diagnosis, prevention, and treatment methods of inflammatory bowel disease. J Med Life. 2019;12:113. 10.25122/jml-2018-0075
37 Mitropoulou M-A, Fradelos EC, Lee KY, Malli F, Tsaras K, Christodoulou NG, et al. Quality of life in patients with inflammatory bowel disease: Importance of psychological symptoms. Cureus. 2022;14:e28502. 10.7759/cureus.28502
38 Curr D, Ianiro G, Pecere S, Bibb S, Cammarota G. Probiotics, fibre and herbal medicinal products for functional and inflammatory bowel disorders. Br J Pharmacol. 2017;174:1426–49. 10.1111/bph.13632
39 Osman N, Adawi D, Molin G, Ahrne S, Berggren A, Jeppsson B. Bifidobacterium infantis strains with and without a combination of oligofructose and inulin (OFI) attenuate inflammation in DSS-induced colitis in rats. BMC Gastroenterol. 2006;6:31. 10.1186/1471-230X-6-31
40 Zhang X, Li M, Wei D, Wang X, Chen X, Xing L. Disruption of the fatty acid Δ6-desaturase gene in the oil-producing fungus Mortierellaisabellina by homologous recombination. Curr Microbiol. 2007;55:128–34. 10.1007/s00284-006-0641-1
41 Allonsius CN, van den Broek MFL, De Boeck I, Kiekens S, Oerlemans EFM, Kiekens F, et al. Interplay between Lactobacillus rhamnosus and Candida and the involvement of exopolysaccharides. Microb Biotechnol. 2017;10:1753–63. 10.1111/1751-7915.12799
42 Bancroft John D, Gamble Marilyn G. Theory and practice of histological techniques, 6th ed. Philadelphia, PA: Churchill Livingstone Elsevier; ©2008, 725 p.
43 Din AU, Hasan A, Zhu Y, Zhang K, Wang Y, Li T, et al. Inhibitory effect of Bifidobacterium bifidum ATCC 29521 on colitis and its mechanism. J Nutr Biochem. 2020;79:108353. 10.1016/j.jnutbio.2020.108353
44 Li D, Chen H, Mao B, Yang Q, Zhao J, Gu Z, et al. Microbial biogeography and core microbiota of the rat digestive tract. Sci Rep. 2017;8:45840. 10.1038/srep45840
45 Das KM. Pharmacotherapy of inflammatory bowel disease. Postgrad Med. 1983;74:141–51. 10.1080/00325481.1983.11698537
46 Plessas S, Nouska C, Karapetsas A, Kazakos S, Alexopoulos A, Mantzourani I, et al. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from Feta-type cheese. Food Chem. 2017;226:102–8. 10.1016/j.foodchem.2017.01.052
47 Arellano K, Vazquez J, Park H, Lim J, Ji Y, Kang HJ, et al. Safety evaluation and whole-genome annotation of Lactobacillus plantarum strains from different sources with special focus on isolates from green tea. Probiotics Antimicrob. Prot. 2020;12:1057–70. 10.1007/s12602-019-09620-y
48 Wang P, Wu Z, Wu J, Pan D, Zeng X, Cheng K. Effects of salt stress on carbohydrate metabolism of Lactobacillus plantarum ATCC 14917. Curr Microbiol. 2016;73:491–7. 10.1007/s00284-016-1087-8
49 Ahn E-M, Kim S-J. The improving effect of gastrodiaelatablume on DSS-induced colitis in mice. Biomed Sci Lett. 2018;24:168–74. 10.15616/BSL.2018.24.3.168
50 Kono Y. Reprint of: generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch Biochem Biophys. 2022;726:109247. 10.1016/j.abb.2022.109247
51 Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–8. 10.1016/0003-2697(79)90738-3
52 Eiserich JP, Hristova M, Cross CE, Jones AD, Freeman BA, Halliwell B, et al. Formation of nitric oxide-derived inflammatory oxidants by myeloperoxidase in neutrophils. Nature. 1998;391:393–7. 10.1038/34923
53 Dejban P, Nikravangolsefid N, Chamanara M, Dehpour A, Rashidian A. The role of medicinal products in the treatment of inflammatory bowel diseases (IBD) through inhibition of TLR4/NF-kappaB pathway. Phytother Res. 2021;35(2):835–45. 10.1002/ptr.6866
54 Muzes G, Molnár B, Tulassay Z, Sipos F. Changes of the cytokine profile in inflammatory bowel diseases. World J Gastroenterol (WJG). 2012;18:5848. 10.3748/wjg.v18.i41.5848
55 Capaldo CT, Farkas AE, Hilgarth RS, Krug SM, Wolf MF, Benedik JK, et al. A. proinflammatory cytokine-induced tight junction remodeling through dynamic self-assembly of claudins. Mol Biol Cell. 2014;25:2710–9. 10.1091/mbc.e14-02-0773
56 Al-Sadi R, Boivin M, Ma T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front Biosci A J Virtual Libr. 2009;14:2765. 10.2741/3413
57 Guo J, Liao M, Wang J. TLR4 signaling in the development of colitis-associated cancer and its possible interplay with microRNA-155. Cell Commun Signal. 2021;19:90. 10.1186/s12964-021-00771-6
58 Pisani A, Rausch P, Bang C, Ellul S, Tabone T, Marantidis Cordina C, et al. Dysbiosis in the gut microbiota in patients with inflammatory bowel disease during remission. Microbiol Spectr. 2022;10:e00616-22. 10.1128/spectrum.00616-22
59 Kim D-Y, Lee T-S, Jung D-H, Song E-J, Jang A-R, Park J-Y, et al. Oral administration of Lactobacillus sakei CVL-001 improves recovery from dextran sulfate sodium-induced colitis in mice by microbiota modulation. Microorganisms. 2023;11:1359. 10.3390/microorganisms11051359
60 Baldelli V, Scaldaferri F, Putignani L, Del Chierico F. The role of Enterobacteriaceae in gut microbiota dysbiosis in inflammatory bowel diseases. Microorganisms. 2021;9:697. 10.3390/microorganisms9040697
61 Chassaing B, Aitken JD, Malleshappa M, Vijay-Kumar M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr Protoc Immunol. 2014;104:15–25. 10.1002/0471142735.im1525s104
62 Choi S, Woo J-K, Jang Y-S, Kang J-H, Jang J-E, Yi T-H, et al. Fermented Pueraria lobata extract ameliorates dextran sulfate sodium-induced colitis by reducing pro-inflammatory cytokines and recovering intestinal barrier function. Lab Anim Res. 2016;32:151–9. 10.5625/lar.2016.32.3.151
63 Ahn E-M, Kim S-J. The improving effect of gastrodiaelatablume on DSS-induced colitis in mice. Biomed Sci Lett. 2018;24:168–74. 10.15616/BSL.2018.24.3.168
64 Silva JP, Navegantes-Lima KC, Oliveira AL, Rodrigues DV, Gaspar SL, Monteiro VV, et al. Protective mechanisms of butyrate on inflammatory bowel disease. Curr Pharm Des. 2018;24:4154–66. 10.2174/1381612824666181001153605
65 Wang Y, Xie Q, Zhang Y, Ma W, Ning K, Xiang J-Y, et al. Combination of probiotics with different functions alleviate DSS-induced colitis by regulating intestinal microbiota, IL-10, and barrier function. Appl Microbiol Biotechnol. 2020;104:335–49. 10.1007/s00253-019-10259-6