
Download
REVIEW ARTICLE
Engineered mesenchymal stem cells: A novel approach for Th2-targeted modulation in allergic asthma
Mahdi Tabasheria#, Amir Mahdi Mahdavia#, Forough Parhizkarb, Seyyed Shamsadin Atharib*
aStudent Research Committee, School of Medicine, Zanjan University of Medical Sciences, Zanjan, Iran
bDepartment of Immunology, School of Medicine, Zanjan University of Medical Sciences, Zanjan, Iran
#These authors contributed equally to this work.
Abstract
Asthma is a widespread allergic condition that has impacted around 300 million people globally. There are various classifications of asthma, one of which is based on T-helper2 (Th2) cells, and in this review, we have focused on Th2 high type and how it is caused. In the following sections, we have explored various treatment approaches for asthma, with a particular emphasis on mesenchymal stem cells (MSCs) as a more effective alternative to conventional treatments. MSCs contribute to asthma management through multiple mechanisms, including the secretion of secretomes, soluble factors, and even interactions with other cells, such as dendritic cells and macrophages. However, as explained later in this review, there are challenges associated with MSCs. In response to these limitations, the development of engineered MSCs offers a novel approach. These engineered MSCs are tailored to improve therapeutic efficacy by boosting their homing efficiency, survival rates, and capacity to modulate immune responses. Engineered MSCs are designed with a variety of genes, each enabling distinct mechanisms that contribute to the effective control of asthma. By specifically targeting Th2 cells, these genetically modified MSCs can modulate immune responses, reduce inflammation, and improve airway function, offering a promising therapeutic strategy for management of asthma.
Key words: asthma, engineering of MSC, mesenchymal stem cells, secretome, T-helper2
*Corresponding author: Seyyed Shamsadin Athari, Department of Immunology, School of Medicine, Zanjan University of Medical Sciences, Zanjan, Iran. Email address: [email protected], [email protected]
Received 9 August 2025; Accepted 10 September 2025; Available online 1 November 2025
Copyright: Tabasheri M, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Asthma
Characteristics, classification, and causes
Airway allergic diseases (AAD) are a group of allergic conditions marked by the infiltration of T-helper 2 (Th2) cells, which include allergic rhinitis (AR), allergic asthma, and chronic sinusitis (CRS).1,2
Asthma is a heterogeneous3 and long-term inflammatory disease defined by inflammation of the airways, increased airway hyperresponsiveness (AHR), and structural changes in the airway remodeling. It is characterized by narrowing of the airways, persistent inflammation of the airways and surrounding tissues, increased growth and response of the airway smooth muscles as well as structural changes in the airways. Alterations in the respiratory tract lead to a wide range of symptoms, including intermittent episodes of breathing difficulty, audible whistling sounds during breathing (wheezing), and persistent coughing. These symptoms are intensified by different ecological influences, such as respiratory viral infections, air pollution, and inhaled allergens.5
Asthma is categorized by different approaches, such as the underlying cause (allergic and nonallergic), inflammation type (eosinophilic and non-eosinophilic), severity, and physiological characteristics.6 Asthma can also be classified based on the level of Th2 involvement, dividing it into two groups: T2-high and non-T2-high. Since most new biological treatments target T2 cytokines, we aim to explore the underlying mechanisms of T2 asthma, focusing on relevant cytokines and other molecular targets. The imbalance between T1 and T2 cytokines is a key factor in the development of T2-high asthma. T2-low asthma, associated with factors such as obesity, smoking, and age, is linked to the activation of Th1 and/or Th17 cells.7 Recent studies highlight the role of Th17/regulatory T cells (Treg) imbalance in steroid-resistant, severe, and neutrophilic asthma.7 Quantitative polymerase chain reaction (qPCR) of bronchial biopsies showed higher interleukin-5 (IL-5) and IL-13 mRNA in the Th2-high cluster, compared to Th2-low and controls. The Th2-low group showed no significant difference from the controls.8,9 The higher levels of Th2 cytokines at sites of allergic inflammation reinforce the notion that Th2 cells play a central role in driving atopic diseases.10–12 Mice studies show that allergic airway inflammation occurs with Th2 cell transfer, and not with Th1 cells.13 The main challenge in asthma is the Th1/Th2 imbalance.14 Th2 cells mediate humoral immunity by releasing cytokines such as IL-4 and IL-5. IL-4 promotes Th2 differentiation and the production of Th2-associated cytokines (IL-4, IL-5, and IL-13). IL-4 also inhibits Th1 cell proliferation and enhances mast cell degranulation, exacerbating allergic reactions.3 By modulating the levels of IL-10, a protein that helps to regulate the immune system can help to prevent allergic inflammation in the airways.14 Allergen exposure triggers antigen-presenting cells (APCs) to release IL-10, driving CD4+ T cells (Th0) to differentiate into Th2 cells for an effective immune response.3 A disruption in Th1/Th2 balance can lead to allergic diseases.15
T-helper 2
Origin, transcription factors, and differentiation
The differentiation of naïve CD4 T cells into Th2 cells is a key process for coordinating immune responses against extracellular parasites and is associated with allergic diseases such as asthma. This transformation is mainly induced by the activation of transcription factors GATA (erythroid transcription factor) binding protein 3 (GATA3) and signal transducer and activator of transcription 6 (STAT6), which are activated in response to IL-4 signaling.16 GATA3 is a key regulator of Th2 cell differentiation, playing a critical role in promoting the expression of cytokines associated with Th2 immune response,17,18 modifying the chromatin landscape to facilitate the expression of Th2 cytokine locus in a coordinated manner.19 Alongside GATA3 and STAT6, additional transcription factors, such as cellular musculoaponeurotic fibrosarcoma (c-Maf), JunB proto-oncogene (JunB), and interferon regulatory factor 4 (IRF-4) also contribute importantly to Th2 differentiation by influencing the expression of essential cytokines and receptors.16
Activation and cytokine production
The first stage of naïve cell differentiation is influenced by the cytokine environment, the concentration of antigens, the type of APCs, and the presence of costimulatory molecules.20 When activated, Th2 cells release several cytokines, including IL-4, IL-5, and IL-13, which impact different immune cells and tissues.21 IL-4 and IL-13 are essential for inducing immunoglobulin E (IgE) class switching in B cells, which enhances the production of allergen-specific IgE antibodies. This mechanism is vital for the development of allergic reactions, as IgE attaches to allergens and helps mast cells and basophils recognize them, leading to degranulation and release of inflammatory mediators.22 IL-5 is widely believed to be a key factor in the maturation and differentiation of eosinophils,23 the accumulation of which tends to be at the site of allergic inflammation, and plays a role in the development of bronchial asthma.24 The presence of eosinophils can lead to airway inflammation and damage, which exacerbates asthma symptoms. Eosinophils in the lungs accumulate and are linked to mucus hypersecretion, AHR, and tissue remodeling in Th2-driven asthma.25
Pathophysiological effects in asthma
Various pathological changes in the airways are driven by the cytokines secreted by Th2 cells. For instance, increased mucus production is a result of IL-13 enhancement of goblet cell hyperplasia, which can lead to airway obstruction.26 Additionally, bronchial hyperresponsiveness can be promoted by IL-4 and IL-13,27 which is a characteristic feature of asthma that leads to breathing difficulty and wheezing.28 Th2 cells are responsible for more than just recruiting eosinophils. Other immune cells, such as mast cells and dendritic cells (DCs), are also influenced by them, which amplify inflammatory response.29 This network of interactions creates a feedback loop that prolongs airway inflammation and leads to chronic symptoms in asthma patients.29
Different immunotherapeutic paths
Among various drug treatments for asthma, inhaled corticosteroids (ICS) are the most effective controllers of asthma. ICS is effective in the expression of infectious cytokine genes, such as IL-4, IL-5, and IL-13, but these must be consumed regularly, otherwise the initial symptoms reappear; moreover, it also has adverse effects, such as growth suppression and cataract.30
Another treatment method is allergen immunotherapy (AIT), which treats allergic diseases by imparting increasing doses of relevant allergens to patients. This method modulates asthma by targeting Th2 and promoting T-naïves differentiation into Th1.31 Although it can trigger a systemic reaction, some meta-analyses presented conflicting results on AIT’s effectiveness in preventing new sensitizations, particularly in pediatric populations, highlighting a need for further research to clarify these outcomes. It should be noted that these therapies might lead to systemic reactions. Additionally, the 3- to 5-year treatment duration poses significant challenges, limiting their practical applications in some patients.32
Moreover, there are some novel immunotherapeutic modes of treating this disease, for instance the use of monoclonal antibody-based treatments. These treatments are designed to reduce infection by targeting cytokines.33 For example, mepolizumab works by blocking IL-5,34 while dupilumab targets the receptor for two other cytokines, IL4 and IL13, by focusing on interleukin-4 receptor alpha (IL-4Rα).33 As a new growing cure, stem cells are mentioned,35 which is the focus of this article to examine their mechanism of action in Th2 functioning in controlling of asthma. As a new growing cure, stem cells are mentioned.
Mesenchymal stem cells
Pioneers in regenerative medicine and immune modulation
Mesenchymal stem cells are remarkable cells with the ability to renew themselves and transform into various cell types, such as fat, bone, cartilage, epithelial, and endothelial cells.36–39 MSCs are an invaluable source of regenerative medicine and are sourced from a variety of tissues. These include bone marrow, peripheral blood, inner organs, and adipose tissue. Additionally, they are harvested from neonatal tissues such as the umbilical cord, amniotic membrane, amniotic fluid, and placenta, making them widely accessible for therapeutic applications.40,41 The use of MSC treatments has demonstrated significant potential in managing various conditions, including hematological diseases, graft-versus-host disease, diabetes, multiple sclerosis, Crohn’s disease, ulcerative colitis, lupus, and in patients undergoing kidney transplantation.42 MSCs are crucial for tissue regeneration and immune system regulation.37,38 They possess an incredible ability to regulate the immune system by inhibiting both innate and acquired immune cells. This powerful function is enabled through direct cell-to-cell communication as well as the release of a diverse array of secretory factors, collectively known as secretome, which includes growth factors, cytokines, and chemokines.43 Given their potential to differentiate and their immune-regulatory capabilities, MSCs are emerging as a new therapeutic tool for a wide range of immune and degenerative diseases,44,45 although overcoming certain limitations is important to fully realize the clinical potential of MSCs.46 MSCs have been shown to suppress inflammatory responses and pathological remodeling in asthma models by modulating immune cells, particularly T cells, and reducing the levels of inflammatory cytokines, such as IL-4, IL-5, and IL-13.47–50
Regulating Th2 to ameliorate asthma
Mesenchymal stem cells exert their therapeutic effects through various mechanisms, including mitochondrial donation,51 protection against apoptosis and oxidative stress,52 and the secretion of extracellular vesicles (EVs) along with soluble factor, such as transforming growth factor-β (TGF-β), prostaglandin E2 (PGE2), and indoleamine 2,3-dioxygenase (IDO), effectively transforming the asthma patient’s environment from a proinflammatory state to an anti-inflammatory state.53 MSCs play a crucial role in alleviating AHR and preventing goblet cell metaplasia, both of which are key contributors to the development of asthma. Their therapeutic potential could significantly enhance asthma management strategies.54–56 MSCs can effectively modulate the Th2 immune pathway by balancing Th1 and Th2 responses. They suppress key Th2 cytokines such as IL-4, IL-5, and IL-13, reducing allergic inflammation and AHR.57–62 MSCs can promote the production of anti-inflammatory cytokines such as IL-10 and interferon gamma (IFN-γ), which help to reduce Th2-dominated allergic responses.62 They significantly enhance IL-10 expression while suppressing the expression of surface receptors.63 Furthermore, MSCs can modulate DC maturation64 and macrophage differentiation to suppress Th2 responses.65
Exosomes
Exosomes, cystic vesicles released by MSCs, are packed with messenger RNA (mRNA), microRNA (miRNA), and proteins.66,67 They transfer these materials into target cells, impacting their growth, death, and movement.68 Studies have shown that miRNAs can control gene expression by binding to specific regions of mRNA.69 For example, upregulating MSC-derived exosomal miR-222-3p leads to reduced expression of interferon regulatory factor 2 (IRF2).70 IRF2 is found to promote Th2 cell immune response and inhibit Th1 cell immune response.71–73 Therefore, overexpression of miR-222-3p through engineering enhances the Th1/Th2 cell ratio, which could be useful in controlling asthma.70
Transforming growth factor-β
Mesenchymal stem cells possess an extraordinary capacity to direct the transformation of CD4+ T cells into CD25+Foxp3+regulatory T cells (induced Tregs) by expressing TGF-β, which produce anti-inflammatory cytokines, such as IL-10 and TGF-β that suppress airway inflammation and cause the cycle to continue.60,74,75 This illustrates the remarkable impact of MSCs in regulating immune response.75 This potential is associated with the stimulation of TGF-β66,67 and IFN-γ68 as well as the recruitment of Tregs,69–71 thereby contributing to the balance between Th1 and Th2 cells,68 so that MSCs secreting TGF-β can be helpful in modulating asthma.
Prostaglandin E2
Prostaglandins, such as lipid-derived autacoids, are crucial in modulating the immune system.76 The secretion of PGE2 by MSCs activates the EP2 and EP4 receptors on macrophages, stimulating them to produce and release IL-10, a cytokine with anti-inflammatory properties.77 Studies have shown that stimulation of EP2 and EP4 receptor subtypes promotes monocyte-derived DC maturation.78 However, the exact reason for the necessity of both EP2 and EP4 receptors for this effect remains unknown. It is possible that the two receptors may synergistically increase cyclic adenosine monophosphate (AMP) levels, or they could have non-overlapping actions that are crucial for induction.76 Additionally, the formation of functional heterodimers by these receptors in macrophages is a potential area of interest for further research.77
Indoleamine 2,3-dioxygenase
The expression of IDO is important for MSCs to suppress T-cell proliferation effectively. The varied levels of IDO expression among MSC donors are strongly associated with their capacity to suppress T-cell responses.79,80 IDO works by hindering T-cell proliferation through tryptophan depletion,81 and the production of kynurenine, a tryptophan metabolite that inhibits T-cell proliferation by activating the stress response kinase GCN2.82,83 Furthermore, IDO expression triggers the transformation of naïve CD4+ T-cells into regulatory CD4+CD25+FOXP3+regulatory T-cells.84
Modulate DC functions to suppress Th2 responses
Dendritic cells are crucial in driving the development of allergic diseases by steering naïve T cells toward becoming T helper 2 cells.85 However, MSCs have been demonstrated to impair the maturation and migration of lung DCs to lymph nodes, thereby alleviating Th2-driven inflammation.4 Additionally, MSCs can suppress the activation of both naïve and effector Th2 cells mediated by DCs and reduce the production of chemokines responsible for recruiting Th2 cells to the airways.4 MSC therapy significantly reduces the population of DCs and macrophages that possess antigen-presenting capabilities.63 MSC treatment leads to a reduction in MHCII expression in DCs, which is responsible for presenting antigens to Th2 cells.63
Regulate macrophage differentiation to inhibit Th2 responses
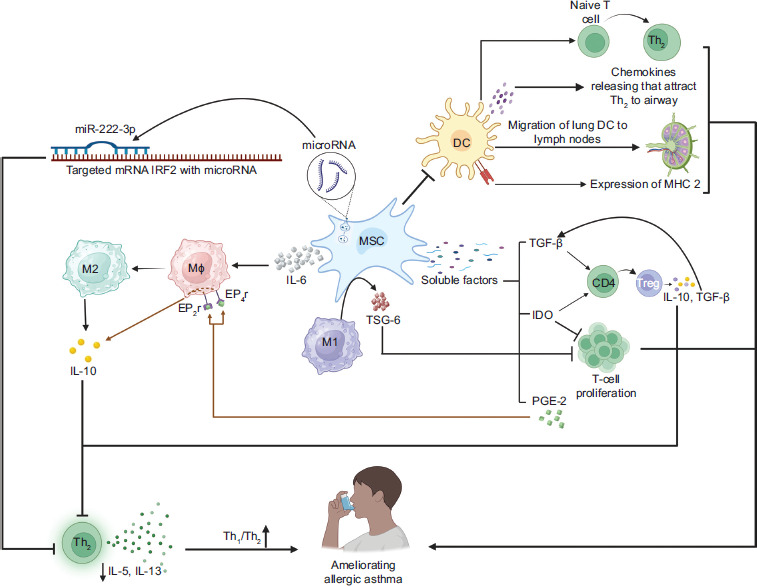
Along with Th2 cells, M2 macrophages are a major source of type 2 cytokines that worsen asthmatic inflammation.86 Macrophages are the primary immune cells found in the airway, able to differentiate into M1 or M2 subtypes depending on particular signals.87 M1 macrophages are primarily induced by pro-inflammatory signals and are characterized by the secretion of IFN-γ, which is derived from Th1 cells, thereby promoting inflammation. In contrast, M2 macrophages are activated by anti-inflammatory signals, such as IL-4 and IL-13, and play a key role in tissue repair and the resolution of inflammation.88 Many studies have discovered a bidirectional effect of direct contact between MSCs and macrophages that affects both cell types and is important for beneficial effects. Direct contact between MSCs and pro-inflammatory macrophages enhances TSG-6 production by MSCs and leads macrophages to M2-like phenotype while suppressing T cell proliferation.89 IL-6, a cytokine secreted by MSCs and present in inflammatory conditions, plays a pivotal role in driving macrophages toward an anti-inflammatory M2-like phenotype, which is characterized by the production of IL-10.90 A summary of this section is provided in Figure 1.
Figure 1 How MSCs exhibit immunosuppressive modulation to induce immune tolerance against asthma. Upregulating miR-222-3p from MSC-derived exosomes reduces the expression of IRF2, which promotes Th2 cell immune responses. Therefore, miR-222-3p expression improves the Th1/Th2 cell ratio in MSCs. The release of soluble factors such as TGF-β, PGE2, and IDO shifts the asthma patient’s environment from pro-inflammatory to anti-inflammatory. MSCs aid in transforming CD4+ T cells into induced Tregs by expressing TGF-β, which produces anti-inflammatory cytokines such as IL-10 and TGF-β, helping balance Th1 and Th2 cells. PGE2 from MSCs activates macrophage EP2 and EP4 receptors, promoting IL-10 production, while IDO suppresses T-cell proliferation and encourages the conversion of naive CD4+ T cells into induced Tregs. MSCs can inhibit DC functions such as directing naive T cells to become Th2 cells, facilitating the maturation and movement of lung DCs to lymph nodes, producing chemokines that attract Th2 cells to the airway, and enhancing MHCII expression for antigen presentation to Th2 cells. Cell contact between MSCs and M1 macrophages enhances TSG-6 production in MSCs, promoting M2-like polarization of macrophages and suppressing T-cell proliferation. Additionally, IL-6 produced by MSCs helps to regulate macrophage polarization toward an IL-10-producing M2-like phenotype.
Mesenchymal stem cells-based therapies
Preclinical trials and limitation
Multiple preclinical studies have shown the effectiveness of MSCs in various animal models of asthma, including ovalbumin (OVA)-induced allergic asthma and house dust mite (HDM)-induced asthma.91–93 Recent animal studies have shown notable outcomes with MSC treatments.57,94–96 Research has demonstrated that MSCs can diminish inflammatory cell infiltration,93 reduce mucus production,93 lessen airway remodeling,97 and enhance lung functioning. The use of chemical agents that simulate in vivo microenvironment, combined with genetic modifications, is studied as a strategy to improve the survival, proliferation, and homing efficiency of MSCs, thereby enhancing their therapeutic effectiveness.98–101 In damaged tissues, the potential for MSC engraftment is limited due to cellular death.102,103 Despite the proven safety of MSCs as a form of cellular treatment, their therapeutic efficacy in human patients is limited in general.104 Decrease in proliferative capacity, pluripotent gene expression, and differentiation potential in MSCs are partially attributed to senescence, which is associated with low telomerase activity.105,106 Engineering of MSC is a strategic approach aimed at enhancing their therapeutic efficacy and overcoming several challenges related to their use. Research has shown that modifying MSCs can significantly improve their beneficial effects. Licensing or pre-stimulation of MSCs, including their transfection with specific genes before application, can significantly enhance their immunomodulatory and tissue-repair capabilities. For instance, cytokine pre-activation with IL-1β, tumor necrosis factor α (TNF-α), or IFN-γ has proved to strengthen MSC-driven repair and the resolution of ventilator-induced lung injury.107 In addition, improving the microenvironment after acid-primed lung injury or using MSCs containing the human IL-10 gene or hepatocyte growth factor (HGF) has shown to lessen the negative effects of untreated MSCs.108 In conclusion, modifying MSCs provides a comprehensive strategy to boost their therapeutic effectiveness for asthma by enhancing their survival, targeting ability, and anti-inflammatory effects.
Anti-inflammatory and immunomodulatory properties
The remarkable therapeutic potential of MSCs can be significantly enhanced through strategic pre-treatment methods, such as exposure to hypoxia, cytokines, and cultivation in a three-dimensional (3D) environment. The priming of MSCs induces a shift in their properties, promoting anti-inflammatory, pro-trophic, and regenerative characteristics, thereby enhancing their overall therapeutic potential.109–115 Furthermore, the dynamic production of both anti- and pro-inflammatory factors by MSCs allows for a fine-tuned regulation of their immunoregulatory properties.116 The conditions of cell culture are crucial in influencing and enhancing the beneficial characteristics of MSCs, highlighting the importance of proper conditioning for improved therapeutic outcomes.110,117,118
Survival enhancement
In the context of cell therapies, it is critical for MSCs to navigate effectively the tissue microenvironments characterized by ischemia, inflammation, and oxidative stress to deliver their therapeutic benefits. However, harsh conditions within these microenvironments often result in low MSC survival rates, with as little as 7% of transplanted MSCs surviving for a few days in infarcted animal myocardium.119 To tackle this challenge, researchers are exploring strategies to boost the survival of transplanted MSCs. Using nonviral gene delivery of pro-survival or anti-apoptotic factors into MSCs is an effective strategy.120,121 Moreover, miRNA delivery provides a different approach by modulating gene expression pathways that support cell survival, eliminating the need for the external expression of growth factors or cytoprotective proteins.122
Improved homing efficiency
Attributed to MSCs’ distinct components, such as cytoskeletal protein filaments (mainly microtubules, actin filaments, and intermediate filaments), along with organized myofilaments and pseudopods around the cell periphery, MSC homing demonstrates strong migratory potential.123 It is a complex process that entails a dynamic interaction of cytokines, chemokines, adhesion molecules, and integrins on the surface of administered MSCs, directing their migration from the bloodstream to damaged tissues.124 To achieve successful transplantation outcomes, MSCs must reach target tissues effectively. Researchers have applied nonviral gene delivery approaches to guide MSCs toward specific tissues or improve their intrinsic tissue-homing ability.125,126 This method holds promise for enhancing the therapeutic effectiveness of MSCs while reducing the required dosage. The homing efficiency of MSCs can be improved by strategically adjusting cell culture conditions, increasing the expression of homing molecules, and employing genetic engineering to modify cell surface receptors. This understanding paves the way for developing more targeted and effective MSC-based therapies for conditions such as asthma, where MSC homing plays a pivotal role in tissue regeneration and immunomodulation.127–130
Optimized gene delivery
Mesenchymal stem cells offer incredible potential in regenerative medicine, and advancements in gene delivery techniques can further enhance their therapeutic benefits. By introducing specific genes, MSCs are tailored to improve survival,121,131 target specific cell membrane receptors,132–134 differentiate into desired cell types,135,136 and produce beneficial growth factors,137,138 cytokines,139,140 and miRNA in exosomes.141,142 Gene delivery is the process of introducing exogenous DNA or RNA into cells to modulate gene expression. Viral gene delivery systems capitalize on the natural efficiency of viruses to encapsulate and transport genetic material into target cells.143 While viral gene delivery systems are highly efficient, they present safety concerns such as the risk of mutations and tumorigenicity.144,145 As such, it’s imperative to explore and develop nonviral gene delivery strategies that not only ensure safety but also maximize efficiency and minimize toxicity.146 Effective nonviral techniques, such as membrane disruption methods for nucleic acid delivery into cells (e.g., microinjection, electroporation, etc.) or the use of nanocarriers for nucleic acid packaging, hold significant potential and require further investigation to fully realize the capabilities of MSCs in regenerative therapies.147
Genetic engineering of MSCs
Mesenchymal stem cells can be engineered into highly effective drug-delivery systems through their modification to enhance their survival and tissue-specific targeting, along with incorporating nonviral gene delivery techniques.148 Furthermore, studies have shown that immunomodulatory factors can be introduced into MSCs for sustained expression, highlighting the versatility of MSCs in reducing inflammation and supporting cell-based therapies.149 Modifying the genetic makeup of MSCs empowers them to produce specific factors that can shield them from cell death, enhance their survival in low-oxygen environments, and improve their natural abilities, such as movement and specialization into particular cell types. Genetic engineering of MSCs is typically carried out using viral vectors, although there is a growing use of nonviral vectors for this purpose.150 Genetic engineering of MSCs can be achieved through the overexpression or suppression of specific genes by using advanced gene-editing approaches, including knock-in and knock-out strategies such as the clustered regularly interspaced short palindromic repeats-associated protein 9 (CRISPR/Cas9) system and RNA silencing techniques. These methods enable precise control of native MSC gene activity or the introduction of exogenous genes for specific therapeutic purposes.151
Interleukin-10
IL-10 is a powerful anti-inflammatory cytokine produced naturally.152 Its remarkable abilities include the capacity to protect and mitigate diseases by staunching inflammation and upholding self-tolerance. This cytokine has demonstrated its therapeutic effects in immune-related disorders, including psoriasis, rheumatoid arthritis, and allergic asthma.153–156 It is clear that MSCs, with their immunomodulatory potential, can be further empowered through IL-10 gene expression to manage effectively the pathophysiology of allergic asthma.14 Recent studies have indicated that IL-10 can suppress Th2 cells, significantly decreasing their production of IL-4 and IL-5.157 The manipulation of MSCs as immunomodulatory cells using immunosuppressive biofactors such as IL-10 could very well emerge as the primary method for not only controlling but also curing asthma.14
Interleukin-35
IL-35 is shown to have an impressive ability to inhibit the growth of CD4+ T cells and elevate the presence of Treg cells, thereby potentially reducing the risk of autoimmune and inflammatory diseases.158 Furthermore, IL-35 can effectively suppress the activity of Th1, Th2, and Th17.158 It is important to highlight that Treg cells, whose proliferation and functioning are enhanced by IL-35, play a key role in suppressing the functioning of other T cells to regulate immune response.159,160 In a study, MSCs were successfully transduced using the pUNO1-mIL35 elastic vector, leading to the production of IL-35 in modified cells.4 Their findings revealed that MSCs transfected with the IL-35 gene exhibited a remarkable ability to control effectively allergic asthma symptoms, including AHR, inflammation of the airways, difficulty breathing, wheezing, and persistent coughing.4,5,41 This discovery holds immense promise for advancing the treatment of allergic asthma and underscores the potential of IL-35 gene expression in enhancing therapeutic outcomes.41
MicroRNA-138-5p (miR-138-5p)
MiR-138-5p has a significant role in the pathophysiology of asthma.161 It is shown that miR-138-5p is involved in various mechanisms that contribute to airway inflammation and hyperresponsiveness, the defining characteristics of asthma.162 The presence of pro-inflammatory cytokines, such as TNF-α and IL-6, reduces inflammatory responses in human MSCs (hMSCs) when miR-138-5p is inhibited.163 This inhibition leads to a reduction in allergic symptoms in mice models of asthma, suggesting that targeting miR-138-5p could be a promising means to manage asthma and improve treatment outcomes.161,163 MiR-138-5p has emerged as a key regulator of deacetylase sirtuin 1 (SIRT1), with its elevated levels being strongly linked to a diverse range of human malignancies.164–166 Notably, the relationship between miR-138-5p and SIRT1 has been associated with pancreatic cancer, highlighting its crucial role in the disease.164 Consider engineering of MSCs with the miR-138-5p gene, a method that offers a well-established set of targets. This approach is anticipated to accelerate the integration of innovative hMSC-based therapy. A recent study has discovered that hMSCs significantly decreased the levels of IL-4, IL-6, IL-5, IL-13, TNF-α, and GATA-3. Notably, hMSCs modified with an miR-138-5p inhibitor exhibited an even greater decrease in these factors. This highlights the potential of this approach for enhanced therapeutic outcomes.163
Erythropoietin (EPO)
The latest research has confirmed that EPO has remarkable antioxidant and anti-inflammatory properties that serve an essential function in enhancing the proliferation and migration of MSCs.167 Furthermore, EPO has been shown to effectively mitigate prolonged histopathological transformations in the lungs of asthma patients.168 It is essential to note that EPO also suppresses the synthesis of IL-4 and IL-5, both of which are pivotal for developing asthma.169 A study conducted in 2018 employed MSCs engineered to express EPO in a mouse model of chronic asthma triggered by OVA.167 The results revealed that both MSCs and EPO–MSCs significantly reduced IL-5, IL-4, and IL-13 levels, with EPO–MSCs showing a stronger inhibitory effect. EPO–MSCs also considerably inhibited the TGF-β1-TAK1-p38MAPK pathway, effectively reducing airway inflammation, mucus secretion, AHR, and airway remodeling.
Indoleamine 2,3-dioxygenase
Indoleamine 2,3-dioxygenase degrades tryptophan, an essential amino acid vital for T-cell metabolism, thereby directly inhibiting T-cell activation and proliferation while promoting T-cell inactivation, apoptosis, and differentiation into Treg cells.170–172 IDO-transfected MSCs enhance the antigen-specific immunosuppressive functioning of Treg cells.173 They also stimulate Treg cells to secrete IL-10 and TGF-β1.174 These findings show that IDO–MSCs not only suppress CD4+ T cell proliferation but also promote Treg cell induction, which helps to lessen asthma by balancing Th1 and Th2 cell activity.7,68,173
Soluble suppression of tumorigenicity 2 (sST2)
sST2 functions as an IL-33 receptor antagonist, which triggers Th2 cytokine production. In a model of ammonium persulfate-induced occupational asthma, human-AT-MSCs overexpressing soluble ST2 effectively inhibit IL-33 production, reduce IgE levels, and significantly alleviate neutrophilic inflammation and airway remodeling. This significant advancement shows great potential for revolutionizing the management of occupational asthma.175 A comprehensive summary of engineering of MSC is presented in Table 1.
Table 1 Genetic engineering of mesenchymal stem cells for asthma treatment: a summary of experimental studies.
| Author (Ref.) | Gene engineered | Vector | Animal model | Outcome on Th2 | Outcome on asthma |
|---|---|---|---|---|---|
| Hou et al.14 | IL-10 | Adenovirus | male BALB/c mice | Reducingpro-inflammatorycytokines,suchasIL-4,IL-5,andIL-13 | Reduction in bronchial and perivascularinflammation,goblet cell hyperplasia, and mucus secretion.Reducing AHR |
| Bao et al.41 | IL-35 | Adenovirus | MaleBALB/c mice | inhibits the growth of CD4+Tcells | The suppression of Th1 and Th2 respectively reduces inflammatoryandallergicdiseases |
| Tang et al.163 | miR-138-5p | Lipofectamine(plasmid vector) | FemaleBALB/C mice | ReductionofIL-4,IL-6,IL-5,IL-13,TNF-α,andGATA-3 | Reduces the number of inflammatorycells,includingeosinophils, lymphocytes, and neutrophils |
| Han et al.167 | EPO | Lentivirus | Female BALB/C mice | Suppresses the synthesis ofIL-4andIL-5 | Effectively suppressed airwayinflammation,mucussecretion, AHR, and airway remodeling |
| Martínez-González et al.175 | sST2 | Lentiviral | Male BALB/c mice | PreventsIL-33production and reduce IgE levels | Alleviates airway remodeling |
Advancements in genetic engineering of MSCs offer a compelling path toward revolutionizing the treatment of allergic asthma. Through strategic genetic modifications, such as overexpressing or silencing specific genes, including the introduction of the genes such as IL-10, IL-35, miR-138-5p, EPO, IDO, and sST2, researchers have unlocked the potential to enhance the immunomodulatory and anti-inflammatory properties of MSCs. The resulting engineered MSCs demonstrate unparalleled abilities to effectively control allergic asthma symptoms, reduce inflammation, and regulate immune response. These findings not only underscore the promise of genetic modification of MSCs but also present a compelling case for the development of more potent treatments for allergic asthma and other immune-mediated diseases.
Conclusion
Mesenchymal stem cells are a new therapeutic way of controlling asthma and other allergic diseases by releasing their exosomes and soluble factors. They suppress Th2 responses by modulating the mechanism of macrophages and DCs. However, we are facing a series of limitations in the use of MSCs; in order to overcome this constraint, engineering of MSCs is growing as a new therapeutic technique for controlling asthma, which can be engineered by special genes to overexpress or inhibit the production of interleukins, RNAs, and different molecules. Major new developments in modifying MSCs provide a considerable mode to change how we treat allergic asthma and other immune-related diseases. The strategic genetic modifications of MSCs not only maximize the therapeutic effects of the delivered genes but also have the potential to improve treatment outcomes. Although all these are at a pre-clinical level, with continued research and the expansion of genetic engineering applications, these approaches could develop into more effective and sustainable treatments for asthma. In addition, the combination of transfecting several genes with one another as utilized in different diseases may become an effective therapy for asthma.
Author’s Contribution
Mahdi Tabasheri and Amir Mahdi Mahdavi drafted the manuscript; Forough Parhizkar revised and Seyyed Shamsadin Athari revised and supervised the drafting.
Conflicts of interest
There was no conflict of interest.
Funding
Not applicable.
REFERENCES
1 Zhang Y, Lan F, and ZL. Update on pathomechanisms and treatments in allergic rhinitis. Allergy, 2022;77(11):3309–19. 10.1111/all.15454
2 Shah SA, Kobayashi M. Pathogenesis of chronic rhinosinusitis with nasal polyp and a prominent T2 endotype. Heliyon. 2023;9(9):e19249. 10.1016/j.heliyon.2023.e19249
3 Wu Z, Nasab EM, Arora P, Athari SS. Study effect of probiotics and prebiotics on treatment of OVA-LPS-induced of allergic asthma inflammation and pneumonia by regulating the TLR4/NF-kB signaling pathway. J Transl Med. 2022;20:130. 10.1186/s12967-022-03337-3
4 Choi JY et al. Effects of human adipose tissue-and bone marrow-derived mesenchymal stem cells on airway inflammation and remodeling in a murine model of chronic asthma. Sci Rep. 2022;12(1):12032. 10.1038/s41598-022-16165-8
5 Wenzel SE. Asthma phenotypes: the evolution from clinical to molecular approach. Nat Med. 2012;18:716–25. 10.1038/nm.2678
6 Ma B, Athari SS, Nasab EH, Zhao L. PI3K/AKT/mTOR and TLR4/MyD88/NF-κB signaling inhibitors attenuate pathological mechanisms of allergic asthma. Inflammation. 2021;44(5):1895–907. 10.1007/s10753-021-01466-3
7 Kuruvilla ME, Lee FE-H, Lee GB. Understanding asthma phenotypes, endotypes, and mechanisms of disease. Clin Rev Allergy Immunol. 2019;56(2):219–33. 10.1007/s12016-018-8712-1
8 Ying S, Humbert M, Barkans J, Corrigan CJ, Pfister R, Menz G, et al. Expression of IL-4 and IL-5 mRNA and protein product by CD4þ and CD8þ T cells, eosinophils, and mast cells in bronchial biopsies obtained from atopic and nonatopic (intrinsic) asthmatics. J Immunol. 1997;158:3539–44. 10.4049/jimmunol.158.7.3539
9 Bradding P, Roberts JA, Britten KM, Montefort S, Djukanovic R, Mueller R, et al. Interleukin-4,-5, and-6 and tumor necrosis factor-alpha in normal and asthmatic airways: Evidence for the human mast cell as a source of these cytokines. Am J Respir Cell Mol Biol. 1994;10:471–80. 10.1165/ajrcmb.10.5.8179909
10 Du Q, Meng W, Athari SS, Wang R. The effect of Co-Q10 on allergic rhinitis and allergic asthma. Allergy Asthma Clin Immunol. 2021;17:32. 10.1186/s13223-021-00534-5
11 Yin J, Yan F, Zheng R, Wu X, Athari SS. Immunomodulatory effect of IL-2-induced bone marrow mononuclear cell therapy on control of allergic asthma. Allergol Immunopathol (Madr). 2023;51(1):110–5. 10.15586/aei.v51i1.746
12 Nasaba EM, Atharib SM, Ghafarzadec S, Nasabd A-RM, Athari SS. Immunomodulatory effects of two silymarin isomers ina Balb/c mouse model of allergic asthma. Allergol Immunopathol (Madr). 2020;48(6):646–53. 10.1016/j.aller.2020.01.003
13 Pei W, Zhang Y, Zhu X, Zhao C, Li X, Lü H, et al. Multitargeted Immunomodulatory Therapy for Viral Myocarditis by Engineered Extracellular Vesicles. ACS Nano, 2024; 18(4):2782-2799. 10.1021/acsnano.3c05847
14 Hou C et al. Effect of transduced mesenchymal stem cells with IL-10 gene on control of allergic asthma. Allergol Immunopathol (Madr). 2023;51(2):45–51. 10.15586/aei.v51i2.789
15 Jacobsen EA et al. Eosinophils and asthma. Curr Allergy Asthma Rep. 2007;7:18–26. 10.1007/s11882-007-0026-y
16 Athari SS, Athari SM, Beyzay F, Movassaghi M, Mortaz E, Taghavi M. Critical role of toll-like receptors in pathophysiology of allergic asthma. Eur J Pharmacol. 2017;808:21–7. 10.1016/j.ejphar.2016.11.047
17 Grogan JL, Mohrs M. Harmon B, Lacy DA, Sedat JW, Locksley RM. Early transcription and silencing of cytokine genes underlie polarization of T helper cell subsets. Immunity. 2001;14(3):205–15. 10.1016/S1074-7613(01)00103-0
18 Spilianakis CG, Flavell RA. Long-range intrachromosomal interactions in the T helper type 2 cytokine locus. Nat Immunol. 2004;5:1017–27. 10.1038/ni1115
19 Qian L, Nasab EM, Athari SM, Athari SS. Mitochondria signaling pathways in allergic asthma. J Investig Med. 2022;70(4):863–82. 10.1136/jim-2021-002098
20 Tao X, Wang J, Yan Y, Cheng P, Liu B, Du H and Niu B. Optimal Sca-1-based procedure for purifying mouse adipose-derived mesenchymal stem cells with enhanced proliferative and differentiation potential. Front. Cell Dev. Biol. 2025; 13:1566670. 10.3389/fcell.2025.1566670
21 Barnes PJ. Th2 cytokines and asthma: An introduction. Respir Res. 2001;2:64. 10.1186/rr39
22 Fujita H, Soyka MB, Akdis M et al. Mechanisms of allergen-specific immunotherapy. Clin Transl Allergy. 2012;2:1–8. 10.1186/2045-7022-2-2
23 Taku Kouro KT. IL-5-and eosinophil-mediated inflammation: From discovery to therapy. Int Immunol. 2009;21(12):1303–9. 10.1093/intimm/dxp102
24 Esmaeilzadeh A, Tahmasebi S, Athari SS. Chimeric antigen receptor-T cell therapy: Applications and challenges in treatment of allergy and asthma. Biomed Pharmacother. 2020;123:109685. 10.1016/j.biopha.2019.109685
25 Hulst GV, Christophe FB, Desmet J. Eosinophils as drivers of severe eosinophilic asthma: Endotypes or plasticity? Int J Mol Sci. 2021;22:10150. 10.3390/ijms221810150
26 Hajimohammadi B, Athari SM, Abdollahi M, Vahedi G, Athari SS. Oral administration of acrylamide worsens the inflammatory responses in the airways of asthmatic mice through agitation of oxidative stress in the lungs. Front Immunol. 2020;11:1940. 10.3389/fimmu.2020.01940
27 Wang H, Li B, Sun Y, Ma Q, Feng Y, Jia Y, et al. NIR-II AIE Luminogen-Based Erythrocyte-Like Nanoparticles with Granuloma-Targeting and Self-Oxygenation Characteristics for Combined Phototherapy of Tuberculosis. Advanced Materials, 2024; 36(38):2406143. 10.1002/adma.202406143
28 Bradding P, Andréanne Côté CP, Dahlén S-E, Hallstrand TS, Brightling CE. Airway hyperresponsiveness in asthma: The role of the epithelium. J Allergy Clin Immunol. 2024;153(5):1181–93. 10.1016/j.jaci.2024.02.011
29 Athari SS. Targeting cell signaling in allergic asthma. Signal Transd Target Ther. 2019;4:45. 10.1038/s41392-019-0079-0
30 Athari SS, Athari SM. The importance of eosinophil, platelet and dendritic cell in asthma. Asian Pac J Trop Dis. 2014;4(1):41–7. 10.1016/S2222-1808(14)60413-8
31 Athari SS, Pourpak Z, Folkerts G, Garssen J, Moin M, Adcock IM, Movassaghi M, Ardestani MS, Moazzeni SM, Mortaz E. Conjugated alpha-alumina nanoparticle with vasoactive intestinal peptide as a nano-drug in treatment of allergic asthma in mice. Eur J Pharmacol. 2016;791:811–20. 10.1016/j.ejphar.2016.10.014
32 Tang L, Chen Y, Xiang Q, Xiang J, Tang Y, et al. The association between IL18, FOXP3 and IL13 genes polymorphisms and risk of allergic rhinitis: a meta-analysis. Inflammation Research, 2020; 69(9):911-923. 10.1007/s00011-020-01368-4
33 Shen Q, Chen J, Yang S, Zhang H, Yu H, Wang S, et al. Protection against cigarette smoke-induced chronic obstructive pulmonary disease via activation of the SIRT1/FoxO1 axis by targeting microRNA-132. American journal of translational research, 2024; 16(10):5516–5524. 10.62347/FVQP4019
34 Zhuang H, Chen Q, Wang W, Qu Q, Xu W, Hu Q, et al. The efficacy of polymyxin B in treating stroke-associated pneumonia with carbapenem-resistant Gram-negative bacteria infections: a multicenter real-world study using propensity score matching. Frontiers in Pharmacology, 2025; 16:1413563. 10.3389/fphar.2025.1413563
35 Trzil JE. Feline asthma: Diagnostic and treatment update. Vet Clin North Am Small Anim Pract. 2020;50(2):375–91. 10.1016/j.cvsm.2019.10.002
36 Nauta AJ, Fibbe WE. Immunomodulatory properties of mesenchymal stromal cells. Blood. 2007;110(10):3499–506. 10.1182/blood-2007-02-069716
37 Hipp J, Atala A. Sources of stem cells for regenerative medicine. Stem Cell Rev. 2008;4(1):3–11. 10.1007/s12015-008-9010-8
38 Rasmusson I. Immune modulation by mesenchymal stem cells. Exp Cell Res. 2006;312(12):2169–79. 10.1016/j.yexcr.2006.03.019
39 Charbord P. Bone marrow mesenchymal stem cells: Historical overview and concepts. Hum Gene Ther. 2010;21(9):1045–56. 10.1089/hum.2010.115
40 Weiss ARR, Dahlke MH. Immunomodulation by mesenchymal stem cells (MSCs): Mechanisms of action of living, apoptotic, and dead MSCs. Front Immunol. 2019;10:1191. 10.3389/fimmu.2019.01191
41 Bao XH et al. Immunomodulatory effect of IL-35 gene-transfected mesenchymal stem cells on allergic asthma. Fundam Clin Pharmacol. 2023;37(1):116–24. 10.1111/fcp.12823
42 Gao F et al. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016;7(1):e2062. 10.1038/cddis.2015.327
43 Marofi F, Vahedi G, Biglari A, Esmaeilzadeh A, Athari SS. Mesenchymal stromal/stem cells: A new era in the cell-based targeted gene therapy of cancer. Front Immunol. 2017;8:1770. 10.3389/fimmu.2017.01770
44 Pittenger MF et al. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen Med. 2019;4:22. 10.1038/s41536-019-0083-6
45 Kim HS et al. Clinical trial of human umbilical cord blood-derived stem cells for the treatment of moderate-to-severe atopic dermatitis: Phase I/IIa studies. Stem Cells. 2017;35(1):248–55. 10.1002/stem.2401
46 Pei M. Environmental preconditioning rejuvenates adult stem cells’ proliferation and chondrogenic potential. Biomaterials. 2017;117:10–23. 10.1016/j.biomaterials.2016.11.049
47 Hong GH et al. hMSCs suppress neutrophil-dominant airway inflammation in a murine model of asthma. Exp Mol Med. 2017;49(1):e288. 10.1038/emm.2016.135
48 Sun YQ et al. Human pluripotent stem cell-derived mesenchymal stem cells prevent allergic airway inflammation in mice. Stem Cells. 2012;30(12):2692–9. 10.1002/stem.1241
49 Castro LL et al. Multiple doses of adipose tissue-derived mesenchymal stromal cells induce immunosuppression in experimental asthma. Stem Cells Transl Med. 2020;9(2):250–60. 10.1002/sctm.19-0120
50 Goodwin M et al. Bone marrow-derived mesenchymal stromal cells inhibit Th2-mediated allergic airways inflammation in mice. Stem Cells. 2011;29(7):1137–48. 10.1002/stem.656
51 Ahmad T et al. Miro1 regulates intercellular mitochondrial transport & enhances mesenchymal stem cell rescue efficacy. Embo J. 2014;33(9):994–1010. 10.1002/embj.201386030
52 Zhang W et al. Research progress of extracellular vesicles and exosomes derived from mesenchymal stem cells in the treatment of oxidative stress-related diseases. Front Immunol. 2023;14:1238789. 10.3389/fimmu.2023.1238789
53 Kim SH et al. Immune inflammatory modulation as a potential therapeutic strategy of stem cell therapy for ALS and neurodegenerative diseases. BMB Rep. 2018;51(11):545–6. 10.5483/BMBRep.2018.51.11.255
54 Takeda K et al. Mesenchymal stem cells recruit CCR2(+) monocytes to suppress allergic airway inflammation. J Immunol. 2018;200(4):1261–9. 10.4049/jimmunol.1700562
55 Yadav UC et al. Aldose reductase inhibition prevents metaplasia of airway epithelial cells. PLoS One. 2010;5(12):e14440. 10.1371/journal.pone.0014440
56 Kuperman DA et al. Direct effects of interleukin-13 on epithelial cells cause airway hyperreactivity and mucus overproduction in asthma. Nat Med. 2002;8(8):885–9. 10.1038/nm734
57 Habibian R, Delirezh N, Farshid AA. The effects of bone marrow-derived mesenchymal stem cells on ovalbumin-induced allergic asthma and cytokine responses in mice. Iran J Basic Med Sci. 2018;21(5):483–8. 10.22038/IJBMS.2018.26898.6575
58 Lambrecht BN, Hammad H. The immunology of asthma. Nat Immunol. 2015;16(1):45–56. 10.1038/ni.3049
59 Lankarani KB, Honarvar B, Athari SS. The mechanisms underlying helicobacter pylori-mediated protection against allergic asthma. Tanaffos. 2017;16(4):251–9.
60 Nemeth K et al. Bone marrow stromal cells use TGF-beta to suppress allergic responses in a mouse model of ragweed-induced asthma. Proc Natl Acad Sci USA. 2010;107(12):5652–7. 10.1073/pnas.0910720107
61 Kang SY et al. Immunologic regulatory effects of human umbilical cord blood-derived mesenchymal stem cells in a murine ovalbumin asthma model. Clin Exp Allergy. 2017;47(7):937–45. 10.1111/cea.12920
62 Abreu SC et al. Bone marrow, adipose, and lung tissue-derived murine mesenchymal stromal cells release different mediators and differentially affect airway and lung parenchyma in experimental asthma. Stem Cells Transl Med. 2017;6(6):1557–67. 10.1002/sctm.16-0398
63 Mo Y et al. Intratracheal administration of mesenchymal stem cells modulates lung macrophage polarization and exerts anti-asthmatic effects. Sci Rep. 2022;12(1):11728. 10.1038/s41598-022-14846-y
64 English K, Barry FP, Mahon BP. Murine mesenchymal stem cells suppress dendritic cell migration, maturation and antigen presentation. Immunol Lett. 2008;115(1):50–8. 10.1016/j.imlet.2007.10.002
65 Selmani Z et al. Human leukocyte antigen-G5 secretion by human mesenchymal stem cells is required to suppress T lymphocyte and natural killer function and to induce CD4+CD25highFOXP3+ regulatory T cells. Stem Cells. 2008;26(1):212–22. 10.1634/stemcells.2007-0554
66 Zhang J et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genom Proteom Bioinform (GPB). 2015;13(1):17–24. 10.1016/j.gpb.2015.02.001
67 Mathivanan S, Ji H, Simpson RJ, Exosomes: extracellular organelles important in intercellular communication. J Proteom. 2010;73(10):1907–20. 10.1016/j.jprot.2010.06.006
68 Masoume Athari S, Mehrabi Nasab E, Shamsadin Athari S. Study effect of Ocimum basilicum seeds on mucus production and cytokine gene expression in allergic asthma mice model. Rev Fr d’Allergol. 2018;58(7):489–93. 10.1016/j.reval.2018.08.003
69 Liang ZZ et al. circRNA-miRNA-mRNA regulatory network in human lung cancer: an update. Cancer Cell Int. 2020;20:173. 10.1186/s12935-020-01245-4
70 Yuan Y et al. Mesenchymal stem cell-derived exosomal miRNA-222-3p increases Th1/Th2 ratio and promotes apoptosis of acute myeloid leukemia cells. Anal Cell Pathol (Amst). 2023;2023:4024887. 10.1155/2023/4024887
71 Le Y et al. Adipogenic mesenchymal stromal cells from bone marrow and their hematopoietic supportive role: Towards understanding the permissive marrow microenvironment in acute myeloid leukemia. Stem Cell Rev Rep. 2016;12(2): 235–44. 10.1007/s12015-015-9639-z
72 Diaz de la Guardia R et al. Detailed characterization of mesenchymal stem/stromal cells from a large cohort of AML patients demonstrates a definitive link to treatment outcomes. Stem Cell Rep. 2017;8(6):1573–86. 10.1016/j.stemcr.2017.04.019
73 Taniguchi T et al. IRF family of transcription factors as regulators of host defense. Ann Rev Immunol. 2001;19:623–55. 10.1146/annurev.immunol.19.1.623
74 Kavanagh H, Mahon BP. Allogeneic mesenchymal stem cells prevent allergic airway inflammation by inducing murine regulatory T cells. Allergy. 2011;66(4):523–31. 10.1111/j.1398-9995.2010.02509.x
75 Patel SA et al. Mesenchymal stem cells protect breast cancer cells through regulatory T cells: Role of mesenchymal stem cell-derived TGF-beta. J Immunol. 2010;184(10):5885–94. 10.4049/jimmunol.0903143
76 Hata AN, Breyer RM. Pharmacology and signaling of prostaglandin receptors: Multiple roles in inflammation and immune modulation. Pharmacol Ther. 2004;103(2):147–66. 10.1016/j.pharmthera.2004.06.003
77 Németh K et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E(2)-dependent reprogramming of host macrophages to increase their Interleukin-10 production. Nat Med. 2009;15(1):42–9. 10.1038/nm.1905
78 Kubo S et al. E-prostanoid (EP)2/EP4 receptor-dependent maturation of human monocyte-derived dendritic cells and induction of helper T2 polarization. J Pharmacol Exp Ther. 2004;309(3):1213–20. 10.1124/jpet.103.062646
79 DelaRosa O et al. Requirement of IFN-gamma-mediated indoleamine 2,3-dioxygenase expression in the modulation of lymphocyte proliferation by human adipose-derived stem cells. Tissue Eng A. 2009;15(10):2795–806. 10.1089/ten.tea.2008.0630
80 François M et al. Human MSC suppression correlates with cytokine induction of indoleamine 2,3-dioxygenase and bystander M2 macrophage differentiation. Mol Ther. 2012;20(1):187–95. 10.1038/mt.2011.189
81 Munn DH, Sharma MD, Mellor AL. Ligation of B7-1/B7-2 by human CD4+ T cells triggers indoleamine 2,3-dioxygenase activity in dendritic cells. J Immunol. 2004;172(7):4100–10. 10.4049/jimmunol.172.7.4100
82 Frumento G et al. Tryptophan-derived catabolites are responsible for inhibition of T and natural killer cell proliferation induced by indoleamine 2,3-dioxygenase. J Exp Med. 2002;196(4):459–68. 10.1084/jem.20020121
83 Munn DH et al. GCN2 kinase in T cells mediates proliferative arrest and anergy induction in response to indoleamine 2,3-dioxygenase. Immunity. 2005;22(5):633–42. 10.1016/j.immuni.2005.03.013
84 Fallarino F et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory phenotype in naive T cells. J Immunol. 2006;176(11):6752–61. 10.4049/jimmunol.176.11.6752
85 Peng YQ et al. Mesenchymal stromal cells-derived small extracellular vesicles modulate DC function to suppress Th2 responses via IL-10 in patients with allergic rhinitis. Eur J Immunol. 2022;52(7):1129–40. 10.1002/eji.202149497
86 Guilliams M et al. Establishment and maintenance of the macrophage niche. Immunity. 2020;52(3):434–51. 10.1016/j.immuni.2020.02.015
87 Muraille E, Leo O. Moser M. TH1/TH2 paradigm extended: macrophage polarization as an unappreciated pathogen-driven escape mechanism? Front Immunol. 2014;5:603. 10.3389/fimmu.2014.00603
88 Bosco MC. Macrophage polarization: Reaching across the aisle? J Allergy Clin Immunol. 2019;143(4):1348–50. 10.1016/j.jaci.2018.12.995
89 Li Y et al. Cell-cell contact with proinflammatory macrophages enhances the immunotherapeutic effect of mesenchymal stem cells in two abortion models. Cell Mol Immunol. 2019;16(12):908–20. 10.1038/s41423-019-0204-6
90 Wang X et al. BM-MSCs protect against liver ischemia/reperfusion injury via HO-1 mediated autophagy. Mol Med Rep. 2018;18(2):2253–62. 10.3892/mmr.2018.9207
91 Shin JW et al. Mesenchymal stem cells suppress severe asthma by directly regulating Th2 cells and type 2 innate lymphoid cells. Mol Cells. 2021;44(8):580–90. 10.14348/molcells.2021.0101
92 Braza F et al. Mesenchymal stem cells induce suppressive macrophages through phagocytosis in a mouse model of asthma. Stem Cells. 2016;34(7):1836–45. 10.1002/stem.2344
93 Sun Y-Q et al. Human pluripotent stem cell-derived mesenchymal stem cells prevent allergic airway inflammation in mice. Stem Cells. 2012;30(12):2692–9. 10.1002/stem.1241
94 Dai R et al. Intratracheal administration of adipose derived mesenchymal stem cells alleviates chronic asthma in a mouse model. BMC Pulm Med. 2018;18(1):131. 10.1186/s12890-018-0701-x
95 Kitoko JZ et al. Therapeutic administration of bone marrow-derived mesenchymal stromal cells reduces airway inflammation without up-regulating Tregs in experimental asthma. Clin Exp Allergy. 2018;48(2):205–16. 10.1111/cea.13048
96 de Castro LL et al. Human adipose tissue mesenchymal stromal cells and their extracellular vesicles act differentially on lung mechanics and inflammation in experimental allergic asthma. Stem Cell Res Ther. 2017;8(1):151. 10.1186/s13287-017-0600-8
97 Abreu SC et al. Serum from asthmatic mice potentiates the therapeutic effects of mesenchymal stromal cells in experimental allergic asthma. Stem Cells Transl Med. 2019;8(3):301–12. 10.1002/sctm.18-0056
98 Li D et al. Low levels of TGF-β1 enhance human umbilical cord-derived mesenchymal stem cell fibronectin production and extend survival time in a rat model of lipopolysaccharide-induced acute lung injury. Mol Med Rep. 2016;14(2):1681–92. 10.3892/mmr.2016.5416
99 Nie H et al. IL-1β pretreatment improves the efficacy of mesenchymal stem cells on acute liver failure by enhancing CXCR4 expression. Stem Cells Int. 2020;2020(1):1498315. 10.1155/2020/1498315
100 Moeinabadi-Bidgoli K et al. Genetic modification and preconditioning strategies to enhance functionality of mesenchymal stromal cells: a clinical perspective. Expert Opin Biol Ther. 2023;23(6):461–78. 10.1080/14712598.2023.2205017
101 Huang M, Nasab EM, Athari SS. Immunoregulatory effect of mesenchymal stem cell via mitochondria signaling pathways in allergic asthma. Saudi J Biol Sci. 2021;28(12):6957–62. 10.1016/j.sjbs.2021.07.071
102 Chen L et al. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS One. 2008;3(4):e1886. 10.1371/journal.pone.0001886
103 Han Y et al. The secretion profile of mesenchymal stem cells and potential applications in treating human diseases. Signal Transduct Target Ther. 2022;7(1):92. 10.1038/s41392-022-00932-0
104 Levy O et al. Shattering barriers toward clinically meaningful MSC therapies. Sci Adv. 2020;6(30):eaba6884. 10.1126/sciadv.aba6884
105 Liu L et al. Telomerase deficiency impairs differentiation of mesenchymal stem cells. Exp Cell Res. 2004;294(1):1–8. 10.1016/j.yexcr.2003.10.03110.1006/excr.1995.1056
106 Sachs PC et al. Defining essential stem cell characteristics in adipose-derived stromal cells extracted from distinct anatomical sites. Cell Tissue Res. 2012;349(2):505–15. 10.1007/s00441-012-1423-7
107 Horie S et al. Cytokine pre-activation of cryopreserved xenogeneic-free human mesenchymal stromal cells enhances resolution and repair following ventilator-induced lung injury potentially via a KGF-dependent mechanism. Intensive Care Med Exp. 2020;8(1):8. 10.1186/s40635-020-0295-5
108 Islam D et al. Identification and modulation of microenvironment is crucial for effective mesenchymal stromal cell therapy in acute lung injury. Am J Resp Crit Care Med. 2019;199(10):1214–24. 10.1164/rccm.201802-0356OC
109 Bulati M et al. The immunomodulatory properties of the human amnion-derived mesenchymal stromal/stem cells are induced by INF-γ produced by activated lymphomonocytes and are mediated by cell-to-cell contact and soluble factors. Front Immunol. 2020;11:54. 10.3389/fimmu.2020.00054
110 Bao X-H, Gao F, Athari SS, Wang H. Immunomodulatory effect of IL-35 gene-transfected mesenchymal stem cells on allergic asthma. Fundam Clin Pharmacol. 2023;37(1):116–24. 10.1111/fcp.12823
111 Cunningham CJ, Redondo-Castro E, Allan SM. The therapeutic potential of the mesenchymal stem cell secretome in ischaemic stroke. J Cereb Blood Flow Metab. 2018;38(8):1276–92. 10.1177/0271678X18776802
112 Ferreira JR et al. Mesenchymal stromal cell secretome: Influencing therapeutic potential by cellular pre-conditioning. Front Immunol. 2018;9:2837. 10.3389/fimmu.2018.02837
113 Miceli V et al. Comparative study of the production of soluble factors in human placenta-derived mesenchymal stromal/stem cells grown in adherent conditions or as aggregates in a catheter-like device. Biochem Biophys Res Commun. 2020;522(1):171–6. 10.1016/j.bbrc.2019.11.069
114 Miceli V et al. Comparison of immunosuppressive and angiogenic properties of human amnion-derived mesenchymal stem cells between 2D and 3D culture systems. Stem Cells Int. 2019;2019:7486279. 10.1155/2019/7486279
115 Wang Y et al. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat Immunol. 2014;15(11):1009–16. 10.1038/ni.3002
116 Jiang J, Nasab EM, Athari SM, Athari SS. Effects of vitamin E and selenium on allergic rhinitis and asthma pathophysiology. Resp Physiol Neurobiol. 2021;286:103614. 10.1016/j.resp.2020.103614
117 Müller L et al. Immunomodulatory properties of mesenchymal stromal cells: An update. Front Cell Dev Biol. 2021;9:637725. 10.3389/fcell.2021.637725
118 Song N, Scholtemeijer M, Shah K. Mesenchymal stem cell immunomodulation: Mechanisms and therapeutic potential. Trends Pharmacol Sci. 2020;41(9):653–64. 10.1016/j.tips.2020.06.009
119 Li L et al. How to improve the survival of transplanted mesenchymal stem cell in ischemic heart ? Stem Cells Int. 2016;2016:9682757. 10.1155/2016/9682757
120 Song H et al. Transfection of mesenchymal stem cells with the FGF-2 gene improves their survival under hypoxic conditions. Mol Cells. 2005;19(3):402–7. 10.1016/S1016-8478(23)13186-4
121 Tang YL et al. Improved graft mesenchymal stem cell survival in ischemic heart with a hypoxia-regulated heme oxygenase-1 vector. J Am Coll Cardiol. 2005;46(7):1339–50. 10.1016/j.jacc.2005.05.079
122 Xu J et al. miR-210 over-expression enhances mesenchymal stem cell survival in an oxidative stress environment through antioxidation and c-Met pathway activation. Sci China Life Sci. 2014;57(10):989–97. 10.1007/s11427-014-4725-z
123 Jin XL, & Wang J. Down-Regulation of INSR Restores Th17/Treg Immune Balance and Alleviates Airway Hyperviscosity in Asthmatic Mice via Inactivation of STAT3 Pathway. Discovery Medicine, 2024; 36(181):372-384. 10.24976/Discov.Med.202436181.35
124 Ullah M, Liu DD, Thakor AS. Mesenchymal stromal cell homing: Mechanisms and strategies for improvement. iScience. 2019;15:421–38. 10.1016/j.isci.2019.05.004
125 Mun JY et al. Minicircle microporation-based non-viral gene delivery improved the targeting of mesenchymal stem cells to an injury site. Biomaterials. 2016;101:310–20. 10.1016/j.biomaterials.2016.05.057
126 Levy O et al. mRNA-engineered mesenchymal stem cells for targeted delivery of interleukin-10 to sites of inflammation. Blood. 2013;122(14):e23–32. 10.1182/blood-2013-04-495119
127 Xu T et al. Aryl hydrocarbon receptor protects lungs from cockroach allergen-induced inflammation by modulating mesenchymal stem cells. J Immunol. 2015;195(12):5539–50. 10.4049/jimmunol.1501198
128 Braza F et al. Mesenchymal stem cells induce suppressive macrophages through phagocytosis in a mouse model of asthma. Stem Cells. 2016;34(7):1836–45. 10.1002/stem.2344
129 De Becker A, Riet IV. Homing and migration of mesenchymal stromal cells: How to improve the efficacy of cell therapy? World J Stem Cells. 2016;8(3):73–87. 10.4252/wjsc.v8.i3.73
130 Hou C, Sun F, Liang Y, Nasab EM, Athari SS. Effect of transduced mesenchymal stem cells with IL-10 gene on control of allergic asthma. Allergol Immunopathol (Madr). 2023;51(2):45–51. 10.15586/aei.v51i2.789
131 Ghasemi F, Sarabi P, Athari SS, Esmaeilzadeh A. Therapeutics strategies against cancer stem cell in breast cancer. Int J Biochem Cell Biol. 2019;109:76–81. 10.1016/j.biocel.2019.01.015
132 Cheng Z et al. Targeted migration of mesenchymal stem cells modified with CXCR4 gene to infarcted myocardium improves cardiac performance. Mol Ther. 2008;16(3):571–9. 10.1038/sj.mt.6300374
133 Huang J et al. Genetic modification of mesenchymal stem cells overexpressing CCR1 increases cell viability, migration, engraftment, and capillary density in the injured myocardium. Circ Res. 2010;106(11):1753–62. 10.1161/CIRCRESAHA.109.196030
134 Kumar S, Ponnazhagan S. Bone homing of mesenchymal stem cells by ectopic alpha 4 integrin expression. Faseb J. 2007;21(14):3917–27. 10.1096/fj.07-8275com
135 Tsuchiya H et al. Chondrogenesis enhanced by overexpression of sox9 gene in mouse bone marrow-derived mesenchymal stem cells. Biochem Biophys Res Commun. 2003;301(2):338–43. 10.1016/S0006-291X(02)03026-7
136 Alberton P et al. Conversion of human bone marrow-derived mesenchymal stem cells into tendon progenitor cells by ectopic expression of scleraxis. Stem Cells Dev. 2012;21(6):846–58. 10.1089/scd.2011.0150
137 Beegle JR et al. Preclinical evaluation of mesenchymal stem cells overexpressing VEGF to treat critical limb ischemia. Mol Ther Methods Clin Dev. 2016;3:16053. 10.1038/mtm.2016.53
138 Pollock K et al. Human mesenchymal stem cells genetically engineered to overexpress brain-derived neurotrophic factor improve outcomes in Huntington’s disease mouse models. Mol Ther. 2016;24(5):965–77. 10.1038/mt.2016.12
139 Choi JJ et al. Mesenchymal stem cells overexpressing interleukin-10 attenuate collagen-induced arthritis in mice. Clin Exp Immunol. 2008;153(2):269–76. 10.1111/j.1365-2249.2008.03683.x
140 Nakajima M et al. Mesenchymal stem cells overexpressing interleukin-10 promote neuroprotection in experimental acute ischemic stroke. Mol Ther Methods Clin Dev. 2017;6:102–11. 10.1016/j.omtm.2017.06.005
141 Yu B et al. Exosomes secreted from GATA-4 overexpressing mesenchymal stem cells serve as a reservoir of anti-apoptotic microRNAs for cardioprotection. Int J Cardiol. 2015;182:349–60. 10.1016/j.ijcard.2014.12.043
142 Baglio SR et al. Human bone marrow-and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res Ther. 2015;6(1):127. 10.1186/s13287-015-0116-z
143 Giacca M, Zacchigna S. Virus-mediated gene delivery for human gene therapy. J Control Release. 2012;161(2):377–88. 10.1016/j.jconrel.2012.04.008
144 Barkholt L et al. Risk of tumorigenicity in mesenchymal stromal cell-based therapies--bridging scientific observations and regulatory viewpoints. Cytotherapy. 2013;15(7):753–9. 10.1016/j.jcyt.2013.03.005
145 Wuchter P et al. Standardization of Good Manufacturing Practice-compliant production of bone marrow-derived human mesenchymal stromal cells for immunotherapeutic applications. Cytotherapy. 2015;17(2):128–39. 10.1016/j.jcyt.2014.04.002
146 Yin H. et al. Non-viral vectors for gene-based therapy. Nat Rev Genet. 2014;15(8):541–55. 10.1038/nrg3763
147 Nayerossadat N, Maedeh T, Ali PA. Viral and nonviral delivery systems for gene delivery. Adv Biomed Res. 2012;1:27. 10.4103/2277-9175.98152
148 Hamann A, Nguyen A, Pannier AK. Nucleic acid delivery to mesenchymal stem cells: A review of nonviral methods and applications. J Biol Eng. 2019;13:7. 10.1186/s13036-019-0140-0
149 Petrakis S et al. Gateway-compatible transposon vector to genetically modify human embryonic kidney and adipose-derived stromal cells. Biotechnol J. 2012;7(7):891–7. 10.1002/biot.201100471
150 Park JS et al. Engineering mesenchymal stem cells for regenerative medicine and drug delivery. Methods. 2015;84:3–16. 10.1016/j.ymeth.2015.03.002
151 Varkouhi AK et al. Genetically modified mesenchymal stromal/stem cells: Application in critical illness. Stem Cell Rev Rep. 2020;16(5):812–27. 10.1007/s12015-020-10000-1
152 Moore KW et al. Interleukin-10 and the interleukin-10 receptor. Ann Rev Immunol. 2001;19:683–765. 10.1146/annurev.immunol.19.1.683
153 Mehrabi Nasab E, Athari SM, Motlagh B, Athari SS. Effects of oral administration of Ocimum basilicum on goblet cell hyperplasia and upstream cytokine gene expression in allergic asthma. Rev Franç d’Allergol. 2020;60:64–8. 10.1016/j.reval.2019.02.226
154 McInnes IB et al. IL-10 improves skin disease and modulates endothelial activation and leukocyte effector function in patients with psoriatic arthritis. J Immunol, 2001;167(7):4075–82. 10.4049/jimmunol.167.7.4075
155 Trachsel E et al. Antibody-mediated delivery of IL-10 inhibits the progression of established collagen-induced arthritis. Arthritis Res Ther. 2007;9(1):R9. 10.1186/ar2115
156 Galeazzi M et al. A phase IB clinical trial with Dekavil (F8-IL10), an immunoregulatory ‘armed antibody’ for the treatment of rheumatoid arthritis, used in combination wiIh methotrexate. Isr Med Assoc J. 2014;16(10):666.
157 Kosaka S et al. IL-10 controls Th2-type cytokine production and eosinophil infiltration in a mouse model of allergic airway inflammation. Immunobiology. 2011;216(7):811–20. 10.1016/j.imbio.2010.12.003
158 Amari A et al. In vitro generation of IL-35-expressing human Wharton’s jelly-derived mesenchymal stem cells using lentiviral vector. Iran J Allergy Asthma Immunol. 2015;14(4):416–26.
159 Bardel E et al. Human CD4+ CD25+ Foxp3+ regulatory T cells do not constitutively express IL-35. J Immunol. 2008;181(10):6898–905. 10.4049/jimmunol.181.10.6898
160 Haribhai D et al. A requisite role for induced regulatory T cells in tolerance based on expanding antigen receptor diversity. Immunity. 2011;35(1):109–22. 10.1016/j.immuni.2011.03.029
161 Qiu YY et al. miR-371, miR-138, miR-544, miR-145, and miR-214 could modulate Th1/Th2 balance in asthma through the combinatorial regulation of Runx3. Am J Transl Res. 2017;9(7):3184–99.
162 Wu TD, Brigham EP, McCormack MC. Asthma in the primary care setting. Med Clin North Am. 2019;103(3):435–52. 10.1016/j.mcna.2018.12.004
163 Tang H et al. Protective effect of miR-138-5p inhibition modified human mesenchymal stem cell on ovalbumin-induced allergic rhinitis and asthma syndrome. J Cell Mol Med. 2021;25(11):5038–49. 10.1111/jcmm.16473
164 Tian S et al. miR-138-5p suppresses autophagy in pancreatic cancer by targeting SIRT1. Oncotarget. 2017;8(7):11071–82. 10.18632/oncotarget.14360
165 Zhu J et al. Long non-coding RNA TUG1 promotes cervical cancer progression by regulating the miR-138-5p-SIRT1 axis. Oncotarget. 2017;8(39):65253–64. 10.18632/oncotarget.18224
166 Wang B et al. MiR-138-5p promotes TNF-α-induced apoptosis in human intervertebral disc degeneration by targeting SIRT1 through PTEN/PI3K/Akt signaling. Exp Cell Res. 2016;345(2):199–205. 10.1016/j.yexcr.2016.05.011
167 Han XP et al. EPO modified MSCs can inhibit asthmatic airway remodeling in an animal model. J Cell Biochem. 2018;119(1):1008–16. 10.1002/jcb.26268
168 Wang W et al. Protective effects of recombinant human erythropoietin against pressure overload-induced left ventricular remodeling and premature death in mice. Tohoku J Exp Med. 2011;225(2):131–43. 10.1620/tjem.225.13110.1620/tjem.256.131
169 Strunk T et al. Erythropoietin inhibits cytokine production of neonatal and adult leukocytes. Acta Paed. 2008;97(1):16–20. 10.1111/j.1651-2227.2007.00560.x
170 Chung DJ et al. Indoleamine 2,3-dioxygenase-expressing mature human monocyte-derived dendritic cells expand potent autologous regulatory T cells. Blood. 2009;114(3):555–63. 10.1182/blood-2008-11-191197
171 Jürgens B et al. Interferon-gamma-triggered indoleamine 2,3-dioxygenase competence in human monocyte-derived dendritic cells induces regulatory activity in allogeneic T cells. Blood. 2009;114(15):3235–43. 10.1182/blood-2008-12-195073
172 Katz JBA, Muller J, Prendergast GC. Indoleamine 2,3-dioxygenase in T-cell tolerance and tumoral immune escape. Immunol Rev. 2008;222:206–21. 10.1111/j.1600-065X.2008.00610.x
173 He Y et al. Indoleamine 2,3-dioxgenase transfected mesenchymal stem cells induce kidney allograft tolerance by increasing the production and function of regulatory t cells. Transplantation. 2015;99(9):1829–38. 10.1097/TP.0000000000000856
174 Li J, Liang L, Lyu B, Cai YS, Zuo Y, Su J, et al. Double trouble: The interaction of PM2.5 and O3 on respiratory hospital admissions. Environmental Pollution, 2023; 338:122665. 10.1016/j.envpol.2023.122665
175 Martínez-González I et al. Human mesenchymal stem cells resolve airway inflammation, hyperreactivity, and histopathology in a mouse model of occupational asthma. Stem Cells Dev. 2014;23(19):2352–63. 10.1089/scd.2013.0616