
Download
ORIGINAL ARTICLE
Lower airway microbiome and metabolomic profiles of recurrent wheezing in infants: a case-control study
Jiebin Chen, Sainan Chen, Huiquan Sun, Yuqing Wang *
Department of Respiratory Medicine, Children’s Hospital of Soochow University, Suzhou, Jiangsu, China
Abstract
Objectives: To characterize the microbiome shifts and subsequent metabolite alterations associated with recurrent wheezing (RW) in infants.
Methods: A total of 33 subjects were enrolled in this study, including 18 infants diagnosed with RW, and 15 normal infants as controls. Bronchoalveolar lavage (BAL) fluid was collected from all the subjects. Bacterial DNA was then isolated and analyzed by 16S ribosomal RNA sequencing. In addition, the metabolomic profile of BAL fluid samples was analyzed with mass spectrometry using complementary chromatographic methods. Spearman’s rank correlation analysis was conducted to explore associations between microbial taxa and metabolites.
Results: The study had 21 (63.6%) boys and 12 (36.4%) girls. The mean age was 26.8 ± 4.9 months. Haemophilus (P = 0.003) and Porphyromonas (P = 0.007) genera showed significant difference between the two groups. The metabolites of “starch and sucrose metabolism pathway” and “pentose phosphate pathway” showed significant correlations with the two bacterial genera. For starch and sucrose metabolism pathway, glucose-6-phosphate showed significant positive correlations with Haemophilus (r = 0.44 and P = 0.009) and Porphyromonas (r = 0.45 and P = 0.008). For pentose phosphate pathway, Sedoheptulose 7-phosphate, an intermediate in the pentose phosphate pathway, showed significantly positive correlations with Haemophilus (r = 0.42 and P = 0.02) and Porphyromonas (r = 0.43 and P = 0.01).
Conclusions: Our study provided new evidence that alteration in respiratory tract microbiome could be associated with RW in infants. By elucidating the microbiome and metabolite profile, we identified novel biomarkers potentially useful for personalized management of RW in infants. The future studies should validate the underlying mechanisms in longitudinal cohorts and explore interventions targeting metabolic–microbial crosstalk.
Key words: lower airway, metabolomics, microbiome, recurrent wheezing
*Corresponding author: Yuqing Wang, Department of Respiratory Medicine, Children’s Hospital of Soochow University, Jingde Road No. 303, Suzhou, Jiangsu 215003, China. Email address: [email protected]
Received 3 June 2025; Accepted 1 August 2025; Available online 1 May 2026
Copyright: Chen J, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Recurrent wheezing (RW) in infants is defined as three or more episodes of wheezing within a period of 12 months, typically occurring in children aged <3 years.1 It is a heterogeneous condition characterized by high-pitched expiratory breathing sounds because of narrowed or inflamed small airways. Epidemiologically, RW affects 5–10% of preschool-aged children globally, with higher prevalence in urbanized regions and low-income populations because of environmental exposures.2,3 Unlike transient viral-induced wheezing, this condition persists beyond infancy and is strongly linked to subsequent development of asthma.4 Longitudinal studies indicate that 30% of the affected children progress to asthma by adolescence, contributing to long-term pulmonary dysfunction and increased healthcare costs.5 It is therefore important to elucidate the etiology of RW for risk stratification and early intervention.
Recent etiological studies have highlighted that RW in infants arises from the interplay of environmental triggers, genetic predisposition, and immune dysregulation.6–8 Environmental factors, such as early-life viral infections, air pollution, and microbiome disruptions, may further modulate immune responses, particularly in genetically vulnerable individuals.9,10 Despite extensive research, the etiology of RW in infants remains elusive. Multidisciplinary approaches integrating multi-omics and longitudinal cohorts are essential to unravel causal pathways and identify actionable therapeutic targets.
Bacterial diversity in the respiratory tract is critical for maintaining immune homeostasis, as commensal microbes regulate epithelial barrier integrity, and modulate inflammatory responses and prime adaptive immunity through metabolite-mediated signaling.11 Disruption in microbial equilibrium is implicated in the pathogenesis of allergic diseases and asthma. Reduced diversity and dominance of proteobacterial taxa were reported to be implicated in airway inflammation and hyperreactivity.12,13 Despite these advances, the composition and functional dynamics of the lower airway microbiome in infants with RW remain poorly understood. In this study, we addressed these gaps by characterizing the lower airway microbiome and metabolomics profile in infants with RW using 16S ribosomal RNA (rRNA) sequencing and untargeted metabolomics. We aimed to characterize the microbiome shifts and subsequent metabolite alterations associated with RW in infants.
Methods
Subjects
This is a prospective study approved by the Ethics Review Board of Children’s Hospital of Soochow University (#2023CS038). We included 15 infants diagnosed with RW at our center between January 2018 and December 2022. Written informed consent was obtained from the guardians of all included participants. The following inclusion criteria were adopted: (1) diagnosed with wheezing by an experienced pediatric pulmonology physician; (2) aged between 12 and 36 months; (3) occurrence of wheezing for more than three times in the past year; and (4) no history of administration of inhaled corticosteroids or antibiotics in the past 2 weeks. In addition, 18 age-matched infants who underwent bronchoscopy because of aspiration of a foreign body were recruited as controls. Subjects with known diagnosis of lung disease, dysfunctional breathing, bronchopulmonary dysplasia, cystic fibrosis, or other acute illness were excluded from the study.
Collection of bronchoalveolar lavage (BAL) fluid
Bronchoalveolar lavage fluid was collected under sterile conditions via flexible bronchoscopy performed under general anesthesia to minimize contamination. By following standardized protocols, the bronchoscope was advanced to the subsegmental bronchus, and three aliquots of sterile saline (1–2 mL/kg per aliquot) were instilled and immediately aspirated, with the first aliquot discarded to reduce upper airway contamination. The retrieved BAL fluid was placed on ice and processed within 30 min to preserve microbial integrity. Samples were centrifuged (4°C, 10 min, 500 ×g) to pellet cellular debris, and the supernatant was aliquoted and stored at -80°C until further detection.
Bacterial DNA isolation and 16S rRNA sequencing
For microbiome profiling, DNA was extracted from the pellet using a validated microbial DNA isolation kit (Qiagen DNeasy PowerSoil Pro kit, Qiagen Inc., Hilden, Germany), incorporating mechanical lysis (bead-beating) to disrupt tough microbial cell walls. Extracted DNA underwent 16S rRNA gene amplification (V3–V4 hypervariable regions) via polymerase chain reaction (PCR), followed by high-throughput sequencing (Illumina MiSeq platforms).11 Negative controls (sterile saline processed identically) were included to account for environmental or reagent contamination.
Paired-end reads were assigned to samples using unique barcodes, followed by truncation of barcode and primer sequences. The trimmed reads were merged with Fast Length Adjustment of Short Reads (FLASH, v1.2.7) to generate raw tags. Demultiplexing and subsequent processing were performed in Quantitative Insights into Microbial Ecology (QIIME) (V1.9.1), where sequence errors were corrected via Deficiency of Adenosine Deaminase 2 (DADA2). A phylogenetic tree was constructed to support phylogenetic diversity analyses. Taxonomic classification was conducted using the SILVA database. The alpha diversity index between groups was analyzed using the R software (Wilcoxon test). Beta diversity was evaluated using weighted and unweighted UniFrac distances, visualized through principal coordinates analysis (PCoA) to illustrate sample clustering. Finally, linear discriminant analysis (LDA) effect size (LEfSe) was applied to identify taxonomy exhibiting significant differences both statistically and biologically.
Metabolomics detection and analysis
For untargeted metabolomics analysis, thawed BAL supernatant was extracted using cold methanol acetonitrile water. After centrifugation (14,000 ×g, 15 min, 4°C), the supernatant was analyzed via liquid chromatography tandem with mass spectrometry (LC-MS) using complementary chromatographic methods as reported previously. High-resolution mass spectrometry (HRMS) was performed in both positive and negative ion modes to maximize metabolite coverage. Raw data were processed using the XCMS software for peak detection, alignment, and noise reduction. Metabolites were annotated with spectral libraries (METLIN) and the functional pathways were annotated using Kyoto Encyclopedia of Genes and Genomes (KEGG).14
Statistical analysis
Statistical analysis was performed using the R software (v4.0.2). Characteristics of the study population were analyzed using the Chi-square test and t-test. Differences in bacterial diversity and relative abundance between the two groups were assessed by the Wilcoxon rank-sum test, with a false discovery rate (FDR) of 5%. Metabolomic data were log-transformed prior to analysis. Multivariate analyses were applied to illustrate differences in samples, including orthogonal partial least squares discriminant analysis (OPLS-DA). Spearman’s rank correlation analysis was conducted to explore associations between microbial taxa and metabolites. All P values were two-tailed, and statistical significance was defined as P < 0.05. Specifically, for LEfSe analysis, LDA > 3.0 and P < 0.05 were considered as statistically significant. For metabolomics, differentially expressed metabolites were based on P < 0.05 from Student’s t-test, and variable importance in projection (VIP) > 1 because of OPLS-DA.
Results
Characteristics of study population
A total of 33 subjects were enrolled in this study, including 18 infants with RW, and 15 infants as controls. There were 21 (63.6%) boys and 12 (36.4%) girls. The mean age was 26.8 ± 4.9 months. All children were from Han population. The baseline characteristics of each group are summarized in Table 1. Patients were found to have higher eosinophil count than the controls with statistical significance (P = 0.001). There was no significant difference in terms of age, gender, body mass index (BMI), and blood WBC and neutrophil counts between the two groups.
Table 1 The baseline characteristics of subjects.
| Recurrent wheezing (RW) group (n = 18) | Control group (n = 15) | P | |
|---|---|---|---|
| Age (month) | 25.3 ± 4.6 | 27.5 ± 5.7 | 0.22 |
| Gender (M/F) | 11/7 | 10/5 | 0.97 |
| BMI (kg/m2) | 13.7 ± 2.1 | 14.1 ± 2.3 | 0.61 |
| WBC counts (×109/L) | 6.5 ± 3.5 | 5.7 ± 3.8 | 0.53 |
| Neutrophils (%) | 62.1 ± 18.6 | 71.3 ± 20.4 | 0.19 |
| Eosinophils (%) | 7.5 ± 3.1 | 3.2 ± 2.5 | 0.001 |
Note: BMI: body mass index; WBC: white blood cells.
Diversity in lower airway microbiome
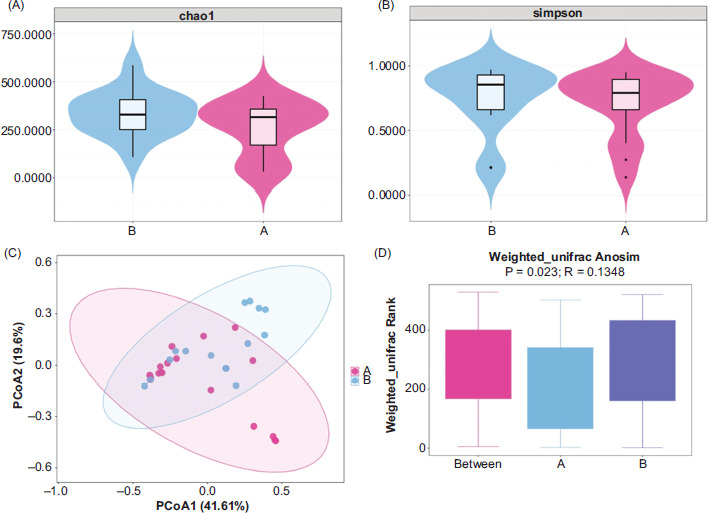
The characteristics of lower airway bacteria in the patients and the controls are shown in Figures 1A and 1B. The Chao1 (richness; P = 0.07) and Simpson (dominance; P = 0.08) indices showed no significant differences between the two groups, which indicated relatively similar bacterial richness and alpha diversity in both groups. The PCoA was performed using weighted UniFrace distance, and Analysis of Similarities (ANOSIM) was performed to evaluate differences between both groups. PCoA and ANOSIM results suggested significant differences in beta diversity between two groups (P = 0.023; Figures 1C and 1D).
Figure 1 Characteristics of the lower airway microbiome. The (A) Chao1 (P = 0.07) and (B) Simpson (P = 0.08) indices indicated similar bacterial richness and alpha diversity in both groups (C) the PCoA plot based on unweighted UniFrac distance and (D) ANOSIM results based on weighted Unifrac showed significant difference in beta diversity between the two groups (P = 0.023).
Distribution of taxa at phylum and genus levels
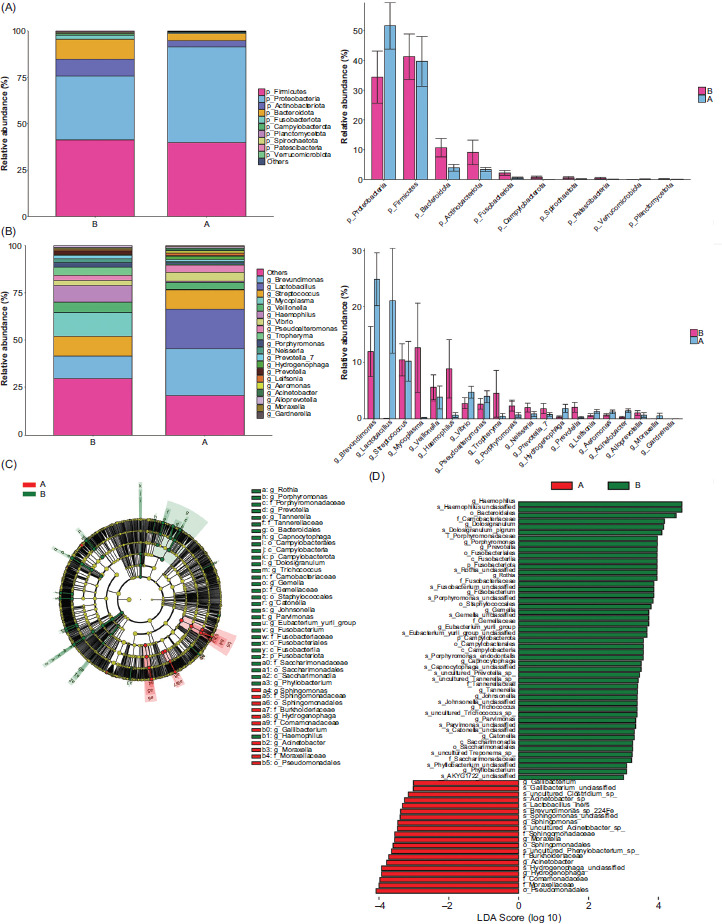
The sequenced data were analyzed according to 35 phyla. The relative abundance of top 10 phyla is shown in Figure 2A. Proteobacteria, Firmicutes, Bacteroidetes, and Actinobacteria were the dominant phyla of bacteria in all samples. Bacteroidota (P = 0.008) and Fusobacteriota (P= 0.01) showed significant difference between the two groups. Figure 2B shows the relative abundance of top 20 genus between the two groups. Haemophilus (P = 0.003) and Porphyromonas (P = 0.007) genera showed significant differences between the two groups.
The LEfSe analysis was performed to identify differences in the composition of bacterial taxa between the two groups. With LDA scores > 3.0, differentially expressed bacteria were identified, some of which were highly abundant in RW infants. Notably, genera Haemophilus and Porphyromonas were significantly abundant in RW infants (P < 0.05; Figures 2C and 2D).
Figure 2 The distribution of taxa at phylum and genus levels. Composition of microbiome at (A) phylum level and (B) genus level. At phylum level, Bacteroidota (P = 0.008) and Fusobacteriota (P=0.01) showed significant difference between the two groups. At genus level, Haemophilus (P = 0.003) and Porphyromonas (P = 0.007) showed significant difference between the two groups. The height of bars represents relative abundance. (C) Bacterial taxa differentially abundant are visualized using a cladogram, with the size of each node indicating relative abundance; (D) with LDA scores > 3.0, the graph of LEfSe distinguishes the microbial communities of each group. Genera Haemophilus and Porphyromonas were significantly abundant in RW infants (P < 0.05).
Characteristics of lower airway metabolomics
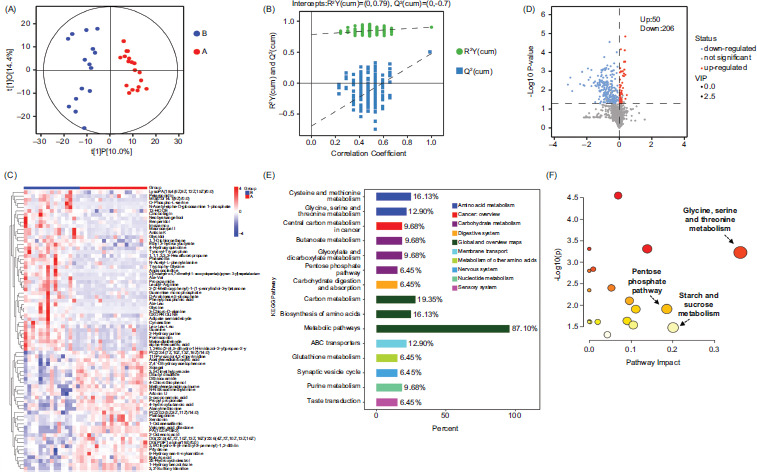
The OPLS-DA result suggested significant differences between the two groups (Figure 3A), with permutation test confirming OPLS-DA model (Figure 3B). Using Student’s t-test (P < 0.05) and VIP > 1 because of OPLS-DA, we identified 256 significantly differentially expressed metabolites, which were predominantly lipids as shown in the heatmap (Figure 3C). Among all the differentially expressed metabolites, 50 were up-regulated and 206 were down-regulated (Figure 3D). Metabolic pathway analyses were then performed to reveal the relationship between differentially expressed metabolites and known metabolic pathways. In all, 20 pathways were significantly impacted in RW infants (Figure 3E). The pathway “glycine, serine and threonine metabolism” was the most significantly impacted in RW infants, followed by “pentose phosphate pathway (PPP)” and “starch and sucrose metabolism pathway” (Figure 3F).
Figure 3 Characteristics of lower airway metabolomics. (A) The OPLS-DA results from all plasma metabolites demonstrated the discriminatory ability of metabolite profiles for both groups; (B) the OPLS-DA model was successfully confirmed with permutation test; (C) heatmap of significant metabolites which differed between the two groups: with higher concentrations, shown in red, and with lower concentrations, shown in blue; (D) volcano plots are displayed, with downregulated and upregulated metabolites shown in blue and red, respectively; (E) KEGG pathway analysis revealed a total of 20 pathways, which were significantly impacted in RW infants; (F) pathway impact represented relative importance of the identified metabolites in the pathway, with larger circles indicating more important pathway impact.
Correlation between lower airway microbiome and metabolites
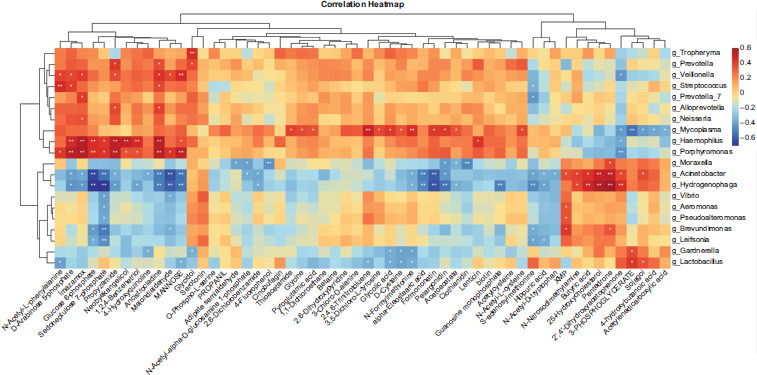
To investigate the relationship between altered microbiome and altered metabolites, we analyzed the correlations between altered genera and altered metabolites using the Spearman’s rank correlation analysis. As summarized in Figure 4, we discovered that the metabolites of starch and sucrose metabolism pathway and PPP showed significant correlations, with the two microbes highly abundant in RW infants (Tables S1 and S2). For starch and sucrose metabolism pathway, glucose-6-phosphate (G6P) showed significantly positive correlations with Haemophilus (r = 0.44 and P = 0.009) and Porphyromonas (r = 0.45 and P = 0.008). For PPP, sedoheptulose 7-phosphate (S7P), an intermediate in the pentose phosphate pathway, showed significantly positive correlations with Haemophilus (r = 0.42 and P = 0.02) and Porphyromonas (r = 0.43 and P = 0.01).
Figure 4 Spearman’s rank correlation analysis for significantly altered metabolites and genera. The metabolites of starch and sucrose metabolism pathway and pentose phosphate pathway (PPP) showed significant correlations with the microbes highly abundant in RW infants. For starch and sucrose metabolism pathway, glucose-6-phosphate (G6P) showed significantly positive correlations with Haemophilus (r = 0.44 and P = 0.009) and Porphyromonas (r = 0.45 and P = 0.008). For PPP, sedoheptulose 7-phosphate (S7P) showed significantly positive correlations with Haemophilus (r = 0.42 and P = 0.02) and Porphyromonas (r = 0.43 and P = 0.01).
Discussion
The pathological mechanism of RW in infants has not been fully elucidated. Recent studies have shown that the human lower airway microbiome could be an etiological factor of various diseases.15–17 Variation of lower airway bacterial diversity is discovered as closely associated with respiratory diseases, such as asthma and cystic fibrosis.18,19 Tang et al.20 investigated the bronchial bacterial microbiome of infants with RW and reported significant differences in diversity in patients, compared to controls. In this study, for the first time, we combined 16S amplicon sequencing and untargeted metabolomics analysis to explore the association of lower airway microbiome with RW. We discovered that RW infants shared some alterations in lower airway microbes and microbiome-derived metabolites. Besides, in line with the findings of Tang et al.,20 we confirmed that RW infants had significantly different lower airway microbiome diversity, which suggested that lower airway microbiome could be associated with the development of RW.
In the current study, we determined two microbes, Haemophilus and Porphyromonas, especially altered in RW infants. Haemophilus was reported to induce neutrophilic inflammation and exacerbate airway hyperreactivity.21 Studies in Haemophilus-colonized murine models demonstrated enhanced airway remodeling driven by transforming growth factor-beta (TGF-β) and matrix metalloproteinases, which could perpetuate wheezing episodes.22 Porphyromonas, a genus traditionally linked to periodontal disease, has recently been implicated in lung diseases. In allergic airways, Porphyromonas gingivalis exacerbates inflammation through activation of interleukin 6 (IL-6) and tumor necrosis factor-α (TNF-α).23 These cytokines may promote neutrophilic infiltration and mucus hypersecretion, which are commonly featured in severe wheezing.24 Taken together, the co-occurrence of these genera could be involved in RW through multiple mechanisms. The potential roles of these distinct microbes in the development of RW need to be further studied.
In addition to microbes, we further investigated the metabolomics profile of lower airway microbiome. We discovered that several pathways were significantly impacted in RW patients. The correlation analysis showed that two metabolites, G6P and S7P, were significantly associated with two microbes identified in microbiome sequencing. G6P and S7P belong to PPP and starch and sucrose metabolism pathway, respectively. G6P is critical for production of nicotinamide adenine dinucleotide phosphate (NADPH) and synthesis of ribose-5-phosphate. G6P-driven glycolysis in epithelial cells may reduce adenosine triphosphate (ATP) available for barrier maintenance.25
In murine asthma models, inhibition of glycolysis reduces airway hyperreactivity.26 S7P is a precursor for ribose-5-phosphate, essential for nucleotide synthesis.27 Excessive S7P could indicate dysregulated PPP activity.28 In hypoxic or inflamed airways, rapid epithelial turnover may increase S7P demand to support DNA repair. Taken together, high G6P with Haemophilus dominance may signal Th17-high inflammation, warranting IL-17 inhibitors. Moreover, elevated S7P with Porphyromonas could indicate mucus hypersecretion, which may suggest anti-inflammation interventions.
The future studies should validate the convergence of G6P and S7P dysregulation with these microbes in longitudinal cohorts. Understanding the underlying mechanisms could pave the way for precision therapies tailored to individual microbial and metabolic profiles.
There were several limitations to this study, which need to be addressed. First, the sample size of the subjects was relatively small, which could limit the power to detect subtle but biologically relevant differences. Recruiting more subjects in the future, study can add to the statistical effectiveness. Second, it cannot be ruled out that the control subjects may have airway inflammation or microbiome disruption, which can potentially lead to selection bias. Third, the cross-sectional nature of the study limited mechanistic investigation. The causal relationship between metabolites and microbial taxa should be further investigated by longitudinal study. Fourth, the taxonomic analysis of our study was limited to the genus level. In the future study, we plan to include shotgun metagenomics sequencing to further explore differences on species level, and if possible, find specific strains of bacteria.
Conclusions
Our study provided new evidence that alteration in the respiratory tract microbiome could be associated with RW in infants. By elucidating the microbiome and metabolite profile, we identified novel biomarkers potentially useful for personalized management of RW in infants. The future studies should validate the underlying mechanisms in longitudinal cohorts and explore interventions targeting metabolic–microbial crosstalk.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that no AI-assisted tools were used in the preparation of this manuscript. All references have been manually verified for accuracy and relevance.
Conflict of Interest
The authors had nothing to disclose regarding funding or any conflict of interest with respect to this manuscript.
Acknowledgment
The authors thanked all the patients who participated in this study.
Author Contributions
All authors contributed equally to this study.
REFERENCES
1 Simões M, Inoue Y, Matsunaga NY, Carvalho MRV, Ribeiro GLT, Morais EO, et al. Recurrent wheezing in preterm infants: Prevalence and risk factors. J Pediatr (Rio J). 2019;95(6):720–7. 10.1016/j.jped.2018.06.007
2 Kumar A, Singh P, Belgrave N. Prevalence of recurrent wheezing in infants and toddlers in Barbados: Findings from a prospective study of a cohort of babies born during 2015–2017. Allergol Immunopathol (Madr). 2021;49(1):17–24. 10.15586/aei.v49i1.21
3 Mallol J, Solé D, Garcia-Marcos L, Rosario N, Aguirre V, Chong H, et al. Prevalence, severity, and treatment of recurrent wheezing during the first year of life: A cross-sectional study of 12,405 Latin American infants. Allergy Asthma Immunol Res. 2016;8(1):22–31. 10.4168/aair.2015.8.1.22
4 Fitzpatrick AM, Bacharier LB, Guilbert TW, Jackson DJ, Szefler SJ, Beigelman A, et al. Phenotypes of recurrent wheezing in preschool children: Identification by latent class analysis and utility in prediction of future exacerbation. J Allergy Clin Immunol Pract. 2019;7(3):915–24.e917. 10.1016/j.jaip.2018.09.016
5 Pijnenburg MW, Frey U, De Jongste JC, Saglani S. Childhood asthma: Pathogenesis and phenotypes. Eur Respir J. 2022;59(6):2100731. 10.1183/13993003.00731-2021
6 Jartti T, Lehtinen P, Vuorinen T, Ruuskanen O. Bronchiolitis: Age and previous wheezing episodes are linked to viral etiology and atopic characteristics. Pediatr Infect Dis J. 2009;28(4):311–7. 10.1097/INF.0b013e31818ee0c1
7 Guo S, Zhang JH, Yan YD, Ding YF, Sheng JY. Association between renin-angiotensin system gene polymorphism and recurrent wheezing in Chinese children: A 4-year follow-up study. J Int Med Res. 2009;37(2):351–8. 10.1177/147323000903700209
8 Korppi M, Nuolivirta K, Lauhkonen E, Holster A, Teräsjärvi J, Vuononvirta J, et al. IL-10 gene polymorphism is associated with preschool atopy and early-life recurrent wheezing after bronchiolitis in infancy. Pediatr Pulmonol. 2017;52(1):14–20. 10.1002/ppul.23489
9 Asharam K, Mitku AAA, Ramsay L, Jeena PM, Naidoo RN. Environmental exposures associated with early childhood recurrent wheezing in the mother and child in the environment birth cohort: A time-to-event study. Thorax. 2024;79(10):953–60. 10.1136/thorax-2023-221150
10 Ayuk AC. Recurrent wheezing and asthma—Epidemiology and environmental determinants. Pediatr Pulmonol. 2024; 60(Suppl. 1):S54–S56. 10.22541/au.172529513.30604546/v1
11 Dickson RP, Erb-Downward JR, Huffnagle GB. Homeostasis and its disruption in the lung microbiome. Am J Physiol Lung Cell Mol Physiol. 2015;309(10):L1047–55. 10.1152/ajplung.00279.2015
12 Losol P, Wolska M, Wypych TP, Yao L, O’Mahony L, Sokolowska M. A cross-talk between microbial metabolites and host immunity: Its relevance for allergic diseases. Clin Transl Allergy 2024;14(2):e12339. 10.1002/clt2.12339
13 Saeed NK, Al-Beltagi M, Bediwy AS, El-Sawaf Y, Toema O. Gut microbiota in various childhood disorders: Implication and indications. World J Gastroenterol. 2022;28(18):1875–1901. 10.3748/wjg.v28.i18.1875
14 Kanehisa M, Goto S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res 2000;28(1):27–30. 10.1093/nar/28.1.27
15 Tangedal S, Nielsen R, Aanerud M, Drengenes C, Husebø GR, Lehmann S, et al. Lower airway microbiota in COPD and healthy controls. Thorax. 2024; 79(3):219–226. 10.1136/thorax-2023-220455
16 Imbert S, Revers M, Enaud R, Orieux A, Camino A, Massri A, et al. Lower airway microbiota compositions differ between influenza, COVID-19 and bacteria-related acute respiratory distress syndromes. Crit Care. 2024;28(1):133. 10.1186/s13054-024-04922-2
17 Campbell S, Gerasimidis K, Milling S, Dicker AJ, Hansen R, Langley RJ. The lower airway microbiome in paediatric health and chronic disease. Paediatr Respir Rev. 2024;52:31–43. 10.1016/j.prrv.2024.02.001
18 Sherrard LJ, Bell SC. Lower airway microbiota for “biomarker” measurements of cystic fibrosis disease progression? Thorax. 2018;73(11):1001–3. 10.1136/thoraxjnl-2018-212165
19 Zhang Q, Cox M, Liang Z, Brinkmann F, Cardenas PA, Duff R, et al. Airway microbiota in severe asthma and relationship to asthma severity and phenotypes. PLoS One. 2016;11(4):e0152724. 10.1371/journal.pone.0152724
20 Tang W, Zhang L, Ai T, Xia W, Xie C, Fan Y, et al. A pilot study exploring the association of bronchial bacterial microbiota and recurrent wheezing in infants with atopy. Front Cell Infect Microbiol. 2023;13:1013809. 10.3389/fcimb.2023.1013809
21 Kim RY, Pinkerton JW, Essilfie AT, Robertson AAB, Baines KJ, Brown AC, et al. Role for NLRP3 inflammasome-mediated, IL-1β-dependent responses in severe, steroid-resistant asthma. Am J Respir Crit Care Med. 2017;196(3):283–97. 10.1164/rccm.201609-1830OC
22 Wang G, Pang Z, Chen-Yu Hsu A, Guan X, Ran N, Yuan Y, et al. Combined treatment with SB203580 and dexamethasone suppresses non-typeable haemophilus influenzae-induced Th17 inflammation response in murine allergic asthma. Eur J Pharmacol. 2019;862:172623. 10.1016/j.ejphar.2019.172623
23 Card JW, Carey MA, Voltz JW, Bradbury JA, Ferguson CD, Cohen EA, et al. Modulation of allergic airway inflammation by the oral pathogen Porphyromonas gingivalis. Infect Immun. 2010;78(6):2488–96. 10.1128/IAI.01270-09
24 Yu X, Cai B, Yu L, Li N, Wu C, Hu Z, et al. Wogonoside ameliorates airway inflammation and mucus hypersecretion via NF-κB/STAT6 signaling in ovalbumin-induced murine acute asthma. J Agric Food Chem. 2024;72(13):7033–42. 10.1021/acs.jafc.3c04082
25 Ma J, Wei K, Liu J, Tang K, Zhang H, Zhu L, et al. Glycogen metabolism regulates macrophage-mediated acute inflammatory responses. Nat Commun. 2020;11(1):1769. 10.1038/s41467-020-15636-8
26 Xuan L, Ren L, Zhang W, Du P, Li B, An Z. Formaldehyde aggravates airway inflammation through induction of glycolysis in an experimental model of asthma exacerbated by lipopolysaccharide. Sci Total Environ. 2024;912:168947. 10.1016/j.scitotenv.2023.168947
27 Nagy C, Haschemi A. Sedoheptulose kinase regulates cellular carbohydrate metabolism by sedoheptulose 7-phosphate supply. Biochem Soc Trans. 2013;41(2):674–80. 10.1042/BST20120354
28 Asamizu S, Xie P, Brumsted CJ, Flatt PM, Mahmud T. Evolutionary divergence of sedoheptulose 7-phosphate cyclases leads to several distinct cyclic products. J Am Chem Soc. 2012;134(29):12219–29. 10.1021/ja3041866
Supplementary
| Data 1 | Data 2 | rho | p value | relation |
|---|---|---|---|---|
| g__Acinetobacter | 1,1−Dichloroethane | −0.341 | 0.053 | negtive |
| g__Lactobacillus | 1,1−Dichloroethane | −0.282 | 0.111 | negtive |
| g__Veillonella | 1,1−Dichloroethane | 0.211 | 0.238 | positive |
| g__Prevotella | 1,1−Dichloroethane | 0.208 | 0.245 | positive |
| g__Prevotella_7 | 1,1−Dichloroethane | 0.191 | 0.286 | positive |
| g__Haemophilus | 1,1−Dichloroethane | 0.157 | 0.383 | positive |
| g__Alloprevotella | 1,1−Dichloroethane | 0.139 | 0.440 | positive |
| g__Hydrogenophaga | 1,1−Dichloroethane | −0.137 | 0.444 | negtive |
| g__Gardnerella | 1,1−Dichloroethane | −0.122 | 0.499 | negtive |
| g__Pseudoalteromonas | 1,1−Dichloroethane | −0.110 | 0.542 | negtive |
| g__Mycoplasma | 1,1−Dichloroethane | 0.108 | 0.549 | positive |
| g__Porphyromonas | 1,1−Dichloroethane | 0.104 | 0.565 | positive |
| g__Aeromonas | 1,1−Dichloroethane | −0.094 | 0.601 | negtive |
| g__Vibrio | 1,1−Dichloroethane | −0.091 | 0.613 | negtive |
| g__Streptococcus | 1,1−Dichloroethane | 0.080 | 0.659 | positive |
| g__Moraxella | 1,1−Dichloroethane | −0.049 | 0.785 | negtive |
| g__Leifsonia | 1,1−Dichloroethane | 0.043 | 0.812 | positive |
| g__Neisseria | 1,1−Dichloroethane | −0.025 | 0.890 | negtive |
| g__Tropheryma | 1,1−Dichloroethane | 0.017 | 0.927 | positive |
| g__Brevundimonas | 1,1−Dichloroethane | −0.010 | 0.956 | negtive |
| g__Haemophilus | 1,2,4−Benzenetriol | 0.458 | 0.007 | positive |
| g__Hydrogenophaga | 1,2,4−Benzenetriol | −0.390 | 0.026 | negtive |
| g__Porphyromonas | 1,2,4−Benzenetriol | 0.351 | 0.045 | positive |
| g__Acinetobacter | 1,2,4−Benzenetriol | −0.323 | 0.068 | negtive |
| g__Veillonella | 1,2,4−Benzenetriol | 0.321 | 0.069 | positive |
| g__Streptococcus | 1,2,4−Benzenetriol | 0.291 | 0.100 | positive |
| g__Brevundimonas | 1,2,4−Benzenetriol | −0.276 | 0.120 | negtive |
| g__Prevotella | 1,2,4−Benzenetriol | 0.274 | 0.123 | positive |
| g__Alloprevotella | 1,2,4−Benzenetriol | 0.263 | 0.139 | positive |
| g__Tropheryma | 1,2,4−Benzenetriol | 0.263 | 0.139 | positive |
| g__Neisseria | 1,2,4−Benzenetriol | 0.255 | 0.151 | positive |
| g__Gardnerella | 1,2,4−Benzenetriol | −0.246 | 0.168 | negtive |
| g__Vibrio | 1,2,4−Benzenetriol | −0.203 | 0.257 | negtive |
| g__Leifsonia | 1,2,4−Benzenetriol | −0.198 | 0.269 | negtive |
| g__Moraxella | 1,2,4−Benzenetriol | −0.178 | 0.321 | negtive |
| g__Mycoplasma | 1,2,4−Benzenetriol | 0.164 | 0.362 | positive |
| g__Prevotella_7 | 1,2,4−Benzenetriol | 0.151 | 0.402 | positive |
| g__Aeromonas | 1,2,4−Benzenetriol | −0.139 | 0.440 | negtive |
| g__Pseudoalteromonas | 1,2,4−Benzenetriol | −0.133 | 0.459 | negtive |
| g__Lactobacillus | 1,2,4−Benzenetriol | −0.118 | 0.512 | negtive |
| g__Mycoplasma | 2,4,6−Trinitrotoluene | 0.483 | 0.004 | positive |
| g__Lactobacillus | 2,4,6−Trinitrotoluene | −0.350 | 0.047 | negtive |
| g__Moraxella | 2,4,6−Trinitrotoluene | −0.319 | 0.070 | negtive |
| g__Porphyromonas | 2,4,6−Trinitrotoluene | 0.270 | 0.129 | positive |
| g__Pseudoalteromonas | 2,4,6−Trinitrotoluene | 0.267 | 0.134 | positive |
| g__Gardnerella | 2,4,6−Trinitrotoluene | −0.259 | 0.146 | negtive |
| g__Haemophilus | 2,4,6−Trinitrotoluene | 0.231 | 0.195 | positive |
| g__Veillonella | 2,4,6−Trinitrotoluene | 0.228 | 0.203 | positive |
| g__Hydrogenophaga | 2,4,6−Trinitrotoluene | −0.226 | 0.205 | negtive |
| g__Vibrio | 2,4,6−Trinitrotoluene | 0.205 | 0.252 | positive |
| g__Prevotella | 2,4,6−Trinitrotoluene | 0.200 | 0.265 | positive |
| g__Aeromonas | 2,4,6−Trinitrotoluene | 0.177 | 0.323 | positive |
| g__Neisseria | 2,4,6−Trinitrotoluene | 0.173 | 0.334 | positive |
| g__Prevotella_7 | 2,4,6−Trinitrotoluene | 0.172 | 0.340 | positive |
| g__Alloprevotella | 2,4,6−Trinitrotoluene | 0.166 | 0.355 | positive |
| g__Acinetobacter | 2,4,6−Trinitrotoluene | −0.161 | 0.370 | negtive |
| g__Brevundimonas | 2,4,6−Trinitrotoluene | 0.085 | 0.636 | positive |
| g__Tropheryma | 2,4,6−Trinitrotoluene | 0.081 | 0.652 | positive |
| g__Leifsonia | 2,4,6−Trinitrotoluene | 0.079 | 0.662 | positive |
| g__Streptococcus | 2,4,6−Trinitrotoluene | −0.010 | 0.958 | negtive |
| g__Porphyromonas | 2’,4’−Dihydroxyacetophenone | −0.450 | 0.009 | negtive |
| g__Hydrogenophaga | 2’,4’−Dihydroxyacetophenone | 0.448 | 0.010 | positive |
| g__Veillonella | 2’,4’−Dihydroxyacetophenone | −0.425 | 0.014 | negtive |
| g__Mycoplasma | 2’,4’−Dihydroxyacetophenone | −0.411 | 0.017 | negtive |
| g__Acinetobacter | 2’,4’−Dihydroxyacetophenone | 0.386 | 0.027 | positive |
| g__Haemophilus | 2’,4’−Dihydroxyacetophenone | −0.374 | 0.032 | negtive |
| g__Lactobacillus | 2’,4’−Dihydroxyacetophenone | 0.336 | 0.057 | positive |
| g__Prevotella | 2’,4’−Dihydroxyacetophenone | −0.282 | 0.112 | negtive |
| g__Alloprevotella | 2’,4’−Dihydroxyacetophenone | −0.271 | 0.127 | negtive |
| g__Tropheryma | 2’,4’−Dihydroxyacetophenone | −0.248 | 0.164 | negtive |
| g__Neisseria | 2’,4’−Dihydroxyacetophenone | −0.240 | 0.178 | negtive |
| g__Gardnerella | 2’,4’−Dihydroxyacetophenone | 0.236 | 0.185 | positive |
| g__Moraxella | 2’,4’−Dihydroxyacetophenone | 0.234 | 0.191 | positive |
| g__Prevotella_7 | 2’,4’−Dihydroxyacetophenone | −0.226 | 0.206 | negtive |
| g__Streptococcus | 2’,4’−Dihydroxyacetophenone | −0.190 | 0.290 | negtive |
| g__Vibrio | 2’,4’−Dihydroxyacetophenone | 0.189 | 0.292 | positive |
| g__Brevundimonas | 2’,4’−Dihydroxyacetophenone | 0.171 | 0.339 | positive |
| g__Aeromonas | 2’,4’−Dihydroxyacetophenone | 0.138 | 0.445 | positive |
| g__Leifsonia | 2’,4’−Dihydroxyacetophenone | 0.098 | 0.589 | positive |
| g__Pseudoalteromonas | 2’,4’−Dihydroxyacetophenone | 0.097 | 0.592 | positive |
| g__Acinetobacter | 2,6−Dichlorobenzamide | −0.373 | 0.033 | negtive |
| g__Hydrogenophaga | 2,6−Dichlorobenzamide | −0.371 | 0.034 | negtive |
| g__Mycoplasma | 2,6−Dichlorobenzamide | 0.273 | 0.124 | positive |
| g__Moraxella | 2,6−Dichlorobenzamide | −0.271 | 0.127 | negtive |
| g__Haemophilus | 2,6−Dichlorobenzamide | 0.236 | 0.186 | positive |
| g__Brevundimonas | 2,6−Dichlorobenzamide | −0.136 | 0.450 | negtive |
| g__Leifsonia | 2,6−Dichlorobenzamide | −0.130 | 0.471 | negtive |
| g__Gardnerella | 2,6−Dichlorobenzamide | −0.120 | 0.507 | negtive |
| g__Veillonella | 2,6−Dichlorobenzamide | −0.086 | 0.633 | negtive |
| g__Prevotella_7 | 2,6−Dichlorobenzamide | −0.082 | 0.648 | negtive |
| g__Porphyromonas | 2,6−Dichlorobenzamide | 0.081 | 0.655 | positive |
| g__Prevotella | 2,6−Dichlorobenzamide | 0.067 | 0.710 | positive |
| g__Streptococcus | 2,6−Dichlorobenzamide | −0.056 | 0.755 | negtive |
| g__Neisseria | 2,6−Dichlorobenzamide | −0.042 | 0.816 | negtive |
| g__Lactobacillus | 2,6−Dichlorobenzamide | −0.038 | 0.834 | negtive |
| g__Pseudoalteromonas | 2,6−Dichlorobenzamide | 0.026 | 0.885 | positive |
| g__Aeromonas | 2,6−Dichlorobenzamide | −0.022 | 0.901 | negtive |
| g__Tropheryma | 2,6−Dichlorobenzamide | −0.017 | 0.924 | negtive |
| g__Vibrio | 2,6−Dichlorobenzamide | 0.016 | 0.928 | positive |
| g__Alloprevotella | 2,6−Dichlorobenzamide | 0.010 | 0.954 | positive |
| g__Mycoplasma | 2,6−Dihydroxypyridine | 0.338 | 0.055 | positive |
| g__Lactobacillus | 2,6−Dihydroxypyridine | −0.244 | 0.171 | negtive |
| g__Veillonella | 2,6−Dihydroxypyridine | 0.209 | 0.242 | positive |
| g__Prevotella | 2,6−Dihydroxypyridine | 0.186 | 0.299 | positive |
| g__Alloprevotella | 2,6−Dihydroxypyridine | 0.172 | 0.339 | positive |
| g__Prevotella_7 | 2,6−Dihydroxypyridine | 0.156 | 0.386 | positive |
| g__Tropheryma | 2,6−Dihydroxypyridine | 0.135 | 0.452 | positive |
| g__Acinetobacter | 2,6−Dihydroxypyridine | −0.132 | 0.462 | negtive |
| g__Porphyromonas | 2,6−Dihydroxypyridine | 0.124 | 0.492 | positive |
| g__Leifsonia | 2,6−Dihydroxypyridine | 0.113 | 0.530 | positive |
| g__Hydrogenophaga | 2,6−Dihydroxypyridine | −0.110 | 0.540 | negtive |
| g__Moraxella | 2,6−Dihydroxypyridine | −0.107 | 0.552 | negtive |
| g__Veillonella | 4−hydroxybutanoic acid | −0.203 | 0.258 | negtive |
| g__Alloprevotella | 4−hydroxybutanoic acid | −0.179 | 0.319 | negtive |
| g__Haemophilus | 4−hydroxybutanoic acid | −0.113 | 0.531 | negtive |
| g__Aeromonas | 4−hydroxybutanoic acid | 0.111 | 0.540 | positive |
| g__Streptococcus | 4−hydroxybutanoic acid | 0.090 | 0.619 | positive |
| g__Prevotella_7 | 4−hydroxybutanoic acid | −0.075 | 0.677 | negtive |
| g__Vibrio | 4−hydroxybutanoic acid | 0.075 | 0.679 | positive |
| g__Prevotella | 4−hydroxybutanoic acid | −0.067 | 0.710 | negtive |
| g__Brevundimonas | 4−hydroxybutanoic acid | 0.067 | 0.712 | positive |
| g__Pseudoalteromonas | 4−hydroxybutanoic acid | 0.048 | 0.791 | positive |
| g__Neisseria | 4−hydroxybutanoic acid | −0.047 | 0.794 | negtive |
| g__Leifsonia | 4−hydroxybutanoic acid | −0.039 | 0.827 | negtive |
| g__Gardnerella | 4−hydroxybutanoic acid | 0.027 | 0.884 | positive |
| g__Tropheryma | 4−hydroxybutanoic acid | 0.005 | 0.977 | positive |
| g__Acinetobacter | 4−Hydroxyquinoline | −0.371 | 0.034 | negtive |
| g__Gardnerella | 4−Hydroxyquinoline | −0.355 | 0.042 | negtive |
| g__Tropheryma | 4−Hydroxyquinoline | 0.301 | 0.089 | positive |
| g__Porphyromonas | 4−Hydroxyquinoline | 0.298 | 0.093 | positive |
| g__Veillonella | 4−Hydroxyquinoline | 0.286 | 0.107 | positive |
| g__Hydrogenophaga | 4−Hydroxyquinoline | −0.267 | 0.132 | negtive |
| g__Haemophilus | 4−Hydroxyquinoline | 0.263 | 0.140 | positive |
| g__Vibrio | 4−Hydroxyquinoline | −0.255 | 0.151 | negtive |
| g__Prevotella | 4−Hydroxyquinoline | 0.244 | 0.171 | positive |
| g__Streptococcus | 4−Hydroxyquinoline | 0.240 | 0.179 | positive |
| g__Brevundimonas | 4−Hydroxyquinoline | −0.238 | 0.182 | negtive |
| g__Leifsonia | 4−Hydroxyquinoline | −0.206 | 0.250 | negtive |
| g__Aeromonas | 4−Hydroxyquinoline | −0.192 | 0.285 | negtive |
| g__Pseudoalteromonas | 4−Hydroxyquinoline | −0.191 | 0.288 | negtive |
| g__Alloprevotella | 4−Hydroxyquinoline | 0.188 | 0.296 | positive |
| g__Moraxella | 4−Hydroxyquinoline | −0.161 | 0.371 | negtive |
| g__Mycoplasma | 4−Hydroxyquinoline | 0.133 | 0.460 | positive |
| g__Lactobacillus | 4−Hydroxyquinoline | −0.126 | 0.483 | negtive |
| g__Neisseria | 4−Hydroxyquinoline | 0.117 | 0.518 | positive |
| g__Prevotella_7 | 4−Hydroxyquinoline | 0.017 | 0.925 | positive |
| g__Mycoplasma | Acetoacetate | 0.352 | 0.045 | positive |
| g__Moraxella | Acetoacetate | −0.350 | 0.046 | negtive |
| g__Hydrogenophaga | Acetoacetate | −0.350 | 0.047 | negtive |
| g__Acinetobacter | Acetoacetate | −0.278 | 0.118 | negtive |
| g__Gardnerella | Acetoacetate | −0.244 | 0.171 | negtive |
| g__Haemophilus | Acetoacetate | 0.234 | 0.190 | positive |
| g__Lactobacillus | Acetoacetate | −0.219 | 0.219 | negtive |
| g__Brevundimonas | Acetoacetate | −0.188 | 0.294 | negtive |
| g__Vibrio | Acetoacetate | −0.163 | 0.365 | negtive |
| g__Leifsonia | Acetoacetate | −0.160 | 0.375 | negtive |
| g__Porphyromonas | Acetoacetate | 0.142 | 0.429 | positive |
| g__Neisseria | Acetoacetate | 0.137 | 0.448 | positive |
| g__Aeromonas | Acetoacetate | −0.136 | 0.449 | negtive |
| g__Tropheryma | Acetoacetate | 0.117 | 0.517 | positive |
| g__Alloprevotella | Acetoacetate | 0.100 | 0.581 | positive |
| g__Pseudoalteromonas | Acetoacetate | −0.094 | 0.605 | negtive |
| g__Veillonella | Acetoacetate | 0.082 | 0.648 | positive |
| g__Prevotella | Acetoacetate | 0.080 | 0.657 | positive |
| g__Streptococcus | Acetoacetate | −0.043 | 0.811 | negtive |
| g__Prevotella_7 | Acetoacetate | 0.004 | 0.984 | positive |
| g__Acinetobacter | Acetylcysteine | −0.298 | 0.092 | negtive |
| g__Hydrogenophaga | Acetylcysteine | −0.297 | 0.093 | negtive |
| g__Prevotella | Acetylcysteine | 0.274 | 0.124 | positive |
| g__Gardnerella | Acetylcysteine | −0.256 | 0.151 | negtive |
| g__Moraxella | Acetylcysteine | −0.247 | 0.167 | negtive |
| g__Mycoplasma | Acetylcysteine | 0.196 | 0.275 | positive |
| g__Haemophilus | Acetylcysteine | 0.189 | 0.293 | positive |
| g__Tropheryma | Acetylcysteine | −0.159 | 0.376 | negtive |
| g__Veillonella | Acetylcysteine | 0.145 | 0.420 | positive |
| g__Porphyromonas | Acetylcysteine | 0.123 | 0.497 | positive |
| g__Alloprevotella | Acetylcysteine | 0.110 | 0.544 | positive |
| g__Brevundimonas | Acetylcysteine | −0.104 | 0.565 | negtive |
| g__Prevotella_7 | Acetylcysteine | 0.090 | 0.618 | positive |
| g__Pseudoalteromonas | Acetylcysteine | 0.050 | 0.783 | positive |
| g__Neisseria | Acetylcysteine | 0.047 | 0.794 | positive |
| g__Lactobacillus | Acetylcysteine | 0.046 | 0.798 | positive |
| g__Vibrio | Acetylcysteine | 0.015 | 0.932 | positive |
| g__Leifsonia | Acetylcysteine | −0.015 | 0.935 | negtive |
| g__Streptococcus | Acetylcysteine | 0.012 | 0.948 | positive |
| g__Aeromonas | Acetylcysteine | −0.002 | 0.993 | negtive |
| g__Mycoplasma | Acetylenedicarboxylic acid | −0.351 | 0.045 | negtive |
| g__Haemophilus | Acetylenedicarboxylic acid | −0.300 | 0.090 | negtive |
| g__Tropheryma | Acetylenedicarboxylic acid | −0.213 | 0.235 | negtive |
| g__Gardnerella | Acetylenedicarboxylic acid | 0.203 | 0.258 | positive |
| g__Prevotella_7 | Acetylenedicarboxylic acid | −0.176 | 0.328 | negtive |
| g__Porphyromonas | Acetylenedicarboxylic acid | −0.135 | 0.454 | negtive |
| g__Prevotella | Acetylenedicarboxylic acid | −0.123 | 0.497 | negtive |
| g__Moraxella | Acetylenedicarboxylic acid | 0.121 | 0.501 | positive |
| g__Pseudoalteromonas | Acetylenedicarboxylic acid | −0.112 | 0.536 | negtive |
| g__Lactobacillus | Acetylenedicarboxylic acid | 0.110 | 0.540 | positive |
| g__Neisseria | Acetylenedicarboxylic acid | −0.101 | 0.574 | negtive |
| g__Vibrio | Acetylenedicarboxylic acid | −0.097 | 0.590 | negtive |
| g__Hydrogenophaga | Acetylenedicarboxylic acid | 0.094 | 0.603 | positive |
| g__Veillonella | Acetylenedicarboxylic acid | −0.089 | 0.624 | negtive |
| g__Acinetobacter | Acetylenedicarboxylic acid | 0.088 | 0.624 | positive |
| g__Aeromonas | Acetylenedicarboxylic acid | −0.084 | 0.642 | negtive |
| g__Alloprevotella | Acetylenedicarboxylic acid | −0.059 | 0.744 | negtive |
| g__Leifsonia | Acetylenedicarboxylic acid | −0.053 | 0.769 | negtive |
| g__Brevundimonas | Acetylenedicarboxylic acid | 0.035 | 0.848 | positive |
| g__Streptococcus | Acetylenedicarboxylic acid | −0.006 | 0.973 | negtive |
| g__Moraxella | Adipate semialdehyde | −0.394 | 0.023 | negtive |
| g__Prevotella | Adipate semialdehyde | 0.186 | 0.301 | positive |
| g__Vibrio | Adipate semialdehyde | −0.179 | 0.318 | negtive |
| g__Brevundimonas | Adipate semialdehyde | −0.164 | 0.360 | negtive |
| g__Mycoplasma | Adipate semialdehyde | 0.159 | 0.377 | positive |
| g__Neisseria | Adipate semialdehyde | 0.153 | 0.395 | positive |
| g__Aeromonas | Adipate semialdehyde | −0.138 | 0.445 | negtive |
| g__Porphyromonas | Adipate semialdehyde | 0.122 | 0.499 | positive |
| g__Pseudoalteromonas | Adipate semialdehyde | −0.110 | 0.542 | negtive |
| g__Alloprevotella | Adipate semialdehyde | 0.106 | 0.559 | positive |
| g__Hydrogenophaga | Adipate semialdehyde | −0.098 | 0.585 | negtive |
| g__Leifsonia | Adipate semialdehyde | −0.095 | 0.598 | negtive |
| g__Veillonella | Adipate semialdehyde | 0.092 | 0.609 | positive |
| g__Acinetobacter | Adipate semialdehyde | −0.084 | 0.643 | negtive |
| g__Gardnerella | Adipate semialdehyde | 0.082 | 0.650 | positive |
| g__Haemophilus | Adipate semialdehyde | 0.064 | 0.725 | positive |
| g__Prevotella_7 | Adipate semialdehyde | −0.062 | 0.732 | negtive |
| g__Lactobacillus | Adipate semialdehyde | 0.047 | 0.793 | positive |
| g__Streptococcus | Adipate semialdehyde | −0.027 | 0.882 | negtive |
| g__Tropheryma | Adipate semialdehyde | 0.002 | 0.990 | positive |
| g__Acinetobacter | alpha−Eleostearic acid | −0.496 | 0.004 | negtive |
| g__Hydrogenophaga | alpha−Eleostearic acid | −0.489 | 0.004 | negtive |
| g__Lactobacillus | alpha−Eleostearic acid | −0.262 | 0.141 | negtive |
| g__Streptococcus | alpha−Eleostearic acid | 0.255 | 0.152 | positive |
| g__Porphyromonas | alpha−Eleostearic acid | 0.253 | 0.155 | positive |
| g__Mycoplasma | alpha−Eleostearic acid | 0.232 | 0.194 | positive |
| g__Moraxella | alpha−Eleostearic acid | −0.218 | 0.223 | negtive |
| g__Veillonella | alpha−Eleostearic acid | 0.217 | 0.225 | positive |
| g__Gardnerella | alpha−Eleostearic acid | −0.204 | 0.255 | negtive |
| g__Haemophilus | alpha−Eleostearic acid | 0.199 | 0.268 | positive |
| g__Brevundimonas | alpha−Eleostearic acid | −0.161 | 0.369 | negtive |
| g__Prevotella | alpha−Eleostearic acid | 0.100 | 0.581 | positive |
| g__Tropheryma | alpha−Eleostearic acid | 0.094 | 0.603 | positive |
| g__Alloprevotella | alpha−Eleostearic acid | 0.077 | 0.668 | positive |
| g__Neisseria | alpha−Eleostearic acid | 0.067 | 0.710 | positive |
| g__Leifsonia | alpha−Eleostearic acid | −0.065 | 0.718 | negtive |
| g__Vibrio | alpha−Eleostearic acid | −0.053 | 0.768 | negtive |
| g__Aeromonas | alpha−Eleostearic acid | −0.022 | 0.903 | negtive |
| g__Pseudoalteromonas | alpha−Eleostearic acid | −0.019 | 0.917 | negtive |
| g__Prevotella_7 | alpha−Eleostearic acid | −0.019 | 0.918 | negtive |
| g__Porphyromonas | Ancistrocladine | 0.542 | 0.001 | positive |
| g__Haemophilus | Ancistrocladine | 0.538 | 0.001 | positive |
| g__Hydrogenophaga | Ancistrocladine | −0.515 | 0.002 | negtive |
| g__Acinetobacter | Ancistrocladine | −0.509 | 0.003 | negtive |
| g__Veillonella | Ancistrocladine | 0.432 | 0.012 | positive |
| g__Alloprevotella | Ancistrocladine | 0.398 | 0.022 | positive |
| g__Streptococcus | Ancistrocladine | 0.379 | 0.030 | positive |
| g__Prevotella | Ancistrocladine | 0.365 | 0.037 | positive |
| g__Neisseria | Ancistrocladine | 0.328 | 0.062 | positive |
| g__Brevundimonas | Ancistrocladine | −0.319 | 0.071 | negtive |
| g__Mycoplasma | Ancistrocladine | 0.286 | 0.107 | positive |
| g__Pseudoalteromonas | Ancistrocladine | −0.284 | 0.110 | negtive |
| g__Aeromonas | Ancistrocladine | −0.277 | 0.119 | negtive |
| g__Prevotella_7 | Ancistrocladine | 0.250 | 0.161 | positive |
| g__Vibrio | Ancistrocladine | −0.244 | 0.171 | negtive |
| g__Leifsonia | Ancistrocladine | −0.209 | 0.243 | negtive |
| g__Tropheryma | Ancistrocladine | 0.149 | 0.407 | positive |
| g__Lactobacillus | Ancistrocladine | −0.095 | 0.598 | negtive |
| g__Moraxella | Ancistrocladine | −0.071 | 0.695 | negtive |
| g__Gardnerella | Ancistrocladine | −0.048 | 0.789 | negtive |
| g__Veillonella | Betaine | 0.231 | 0.196 | positive |
| g__Gardnerella | Betaine | −0.209 | 0.243 | negtive |
| g__Lactobacillus | Betaine | −0.208 | 0.245 | negtive |
| g__Prevotella | Betaine | 0.206 | 0.251 | positive |
| g__Acinetobacter | Betaine | −0.201 | 0.260 | negtive |
| g__Mycoplasma | Betaine | 0.150 | 0.405 | positive |
| g__Hydrogenophaga | Betaine | −0.130 | 0.470 | negtive |
| g__Moraxella | Betaine | −0.122 | 0.500 | negtive |
| g__Alloprevotella | Betaine | 0.119 | 0.509 | positive |
| g__Prevotella_7 | Betaine | 0.116 | 0.520 | positive |
| g__Vibrio | Betaine | −0.092 | 0.611 | negtive |
| g__Porphyromonas | Betaine | 0.091 | 0.616 | positive |
| g__Aeromonas | Betaine | −0.090 | 0.617 | negtive |
| g__Haemophilus | Betaine | 0.070 | 0.699 | positive |
| g__Pseudoalteromonas | Betaine | −0.055 | 0.759 | negtive |
| g__Streptococcus | Betaine | 0.045 | 0.803 | positive |
| g__Tropheryma | Betaine | 0.034 | 0.851 | positive |
| g__Neisseria | Betaine | −0.033 | 0.853 | negtive |
| g__Leifsonia | Betaine | 0.030 | 0.870 | positive |
| g__Brevundimonas | Betaine | −0.028 | 0.875 | negtive |
| g__Mycoplasma | biotin | 0.302 | 0.087 | positive |
| g__Acinetobacter | biotin | −0.293 | 0.098 | negtive |
| g__Haemophilus | biotin | 0.292 | 0.099 | positive |
| g__Porphyromonas | biotin | 0.239 | 0.181 | positive |
| g__Hydrogenophaga | biotin | −0.209 | 0.243 | negtive |
| g__Gardnerella | biotin | −0.191 | 0.287 | negtive |
| g__Leifsonia | biotin | −0.189 | 0.291 | negtive |
| g__Moraxella | biotin | −0.157 | 0.383 | negtive |
| g__Streptococcus | biotin | 0.144 | 0.424 | positive |
| g__Veillonella | biotin | 0.127 | 0.481 | positive |
| g__Brevundimonas | biotin | −0.123 | 0.493 | negtive |
| g__Neisseria | biotin | 0.119 | 0.509 | positive |
| g__Aeromonas | biotin | −0.115 | 0.523 | negtive |
| g__Prevotella | biotin | 0.108 | 0.548 | positive |
| g__Pseudoalteromonas | biotin | −0.093 | 0.605 | negtive |
| g__Vibrio | biotin | −0.092 | 0.609 | negtive |
| g__Lactobacillus | biotin | −0.088 | 0.627 | negtive |
| g__Tropheryma | biotin | 0.072 | 0.689 | positive |
| g__Alloprevotella | biotin | 0.052 | 0.775 | positive |
| g__Prevotella_7 | biotin | −0.005 | 0.976 | negtive |
| g__Hydrogenophaga | Butyric acid | 0.469 | 0.006 | positive |
| g__Acinetobacter | Butyric acid | 0.440 | 0.011 | positive |
| g__Porphyromonas | Butyric acid | −0.294 | 0.096 | negtive |
| g__Brevundimonas | Butyric acid | 0.225 | 0.207 | positive |
| g__Haemophilus | Butyric acid | −0.205 | 0.254 | negtive |
| g__Veillonella | Butyric acid | −0.204 | 0.256 | negtive |
| g__Tropheryma | Butyric acid | 0.178 | 0.322 | positive |
| g__Prevotella | Butyric acid | −0.169 | 0.346 | negtive |
| g__Leifsonia | Butyric acid | 0.168 | 0.350 | positive |
| g__Streptococcus | Butyric acid | −0.159 | 0.377 | negtive |
| g__Vibrio | Butyric acid | 0.141 | 0.434 | positive |
| g__Moraxella | Butyric acid | 0.117 | 0.516 | positive |
| g__Gardnerella | Butyric acid | 0.106 | 0.559 | positive |
| g__Pseudoalteromonas | Butyric acid | 0.095 | 0.598 | positive |
| g__Neisseria | Butyric acid | −0.090 | 0.617 | negtive |
| g__Mycoplasma | Butyric acid | −0.084 | 0.644 | negtive |
| g__Aeromonas | Butyric acid | 0.066 | 0.716 | positive |
| g__Prevotella_7 | Butyric acid | −0.053 | 0.771 | negtive |
| g__Alloprevotella | Butyric acid | −0.034 | 0.849 | negtive |
| g__Lactobacillus | Butyric acid | −0.030 | 0.869 | negtive |
| g__Haemophilus | Cinobufagin | 0.309 | 0.080 | positive |
| g__Tropheryma | Cinobufagin | 0.308 | 0.081 | positive |
| g__Hydrogenophaga | Cinobufagin | −0.220 | 0.217 | negtive |
| g__Brevundimonas | Cinobufagin | −0.214 | 0.231 | negtive |
| g__Leifsonia | Cinobufagin | −0.204 | 0.256 | negtive |
| g__Lactobacillus | Cinobufagin | 0.175 | 0.328 | positive |
| g__Acinetobacter | Cinobufagin | −0.123 | 0.493 | negtive |
| g__Prevotella | Cinobufagin | 0.103 | 0.569 | positive |
| g__Gardnerella | Cinobufagin | −0.095 | 0.597 | negtive |
| g__Vibrio | Cinobufagin | −0.073 | 0.687 | negtive |
| g__Moraxella | Cinobufagin | 0.069 | 0.702 | positive |
| g__Alloprevotella | Cinobufagin | −0.069 | 0.703 | negtive |
| g__Neisseria | Cinobufagin | −0.058 | 0.750 | negtive |
| g__Prevotella_7 | Cinobufagin | −0.040 | 0.826 | negtive |
| g__Veillonella | Cinobufagin | −0.031 | 0.864 | negtive |
| g__Porphyromonas | Cinobufagin | −0.031 | 0.866 | negtive |
| g__Aeromonas | Cinobufagin | −0.019 | 0.917 | negtive |
| g__Mycoplasma | Cinobufagin | −0.009 | 0.959 | negtive |
| g__Pseudoalteromonas | Cinobufagin | −0.005 | 0.976 | negtive |
| g__Streptococcus | Cinobufagin | 0.002 | 0.991 | positive |
| g__Moraxella | Clothianidin | −0.466 | 0.006 | negtive |
| g__Hydrogenophaga | Clothianidin | −0.340 | 0.054 | negtive |
| g__Mycoplasma | Clothianidin | 0.281 | 0.113 | positive |
| g__Acinetobacter | Clothianidin | −0.279 | 0.116 | negtive |
| g__Brevundimonas | Clothianidin | −0.267 | 0.133 | negtive |
| g__Aeromonas | Clothianidin | −0.253 | 0.155 | negtive |
| g__Porphyromonas | Clothianidin | 0.243 | 0.173 | positive |
| g__Haemophilus | Clothianidin | 0.236 | 0.185 | positive |
| g__Gardnerella | Clothianidin | −0.232 | 0.193 | negtive |
| g__Veillonella | Clothianidin | 0.225 | 0.208 | positive |
| g__Vibrio | Clothianidin | −0.219 | 0.222 | negtive |
| g__Alloprevotella | Clothianidin | 0.207 | 0.247 | positive |
| g__Leifsonia | Clothianidin | −0.185 | 0.302 | negtive |
| g__Pseudoalteromonas | Clothianidin | −0.173 | 0.335 | negtive |
| g__Prevotella | Clothianidin | 0.155 | 0.390 | positive |
| g__Lactobacillus | Clothianidin | −0.148 | 0.409 | negtive |
| g__Streptococcus | Clothianidin | −0.084 | 0.641 | negtive |
| g__Neisseria | Clothianidin | 0.072 | 0.691 | positive |
| g__Prevotella_7 | Clothianidin | 0.069 | 0.705 | positive |
| g__Tropheryma | Clothianidin | −0.065 | 0.721 | negtive |
| g__Porphyromonas | D−Arabinose 5−phosphate | 0.524 | 0.002 | positive |
| g__Haemophilus | D−Arabinose 5−phosphate | 0.465 | 0.006 | positive |
| g__Hydrogenophaga | D−Arabinose 5−phosphate | −0.445 | 0.010 | negtive |
| g__Acinetobacter | D−Arabinose 5−phosphate | −0.408 | 0.019 | negtive |
| g__Streptococcus | D−Arabinose 5−phosphate | 0.405 | 0.020 | positive |
| g__Veillonella | D−Arabinose 5−phosphate | 0.387 | 0.026 | positive |
| g__Neisseria | D−Arabinose 5−phosphate | 0.338 | 0.055 | positive |
| g__Prevotella | D−Arabinose 5−phosphate | 0.331 | 0.060 | positive |
| g__Tropheryma | D−Arabinose 5−phosphate | 0.310 | 0.079 | positive |
| g__Alloprevotella | D−Arabinose 5−phosphate | 0.303 | 0.086 | positive |
| g__Mycoplasma | D−Arabinose 5−phosphate | 0.286 | 0.107 | positive |
| g__Moraxella | D−Arabinose 5−phosphate | −0.273 | 0.124 | negtive |
| g__Prevotella_7 | D−Arabinose 5−phosphate | 0.231 | 0.195 | positive |
| g__Lactobacillus | D−Arabinose 5−phosphate | −0.229 | 0.200 | negtive |
| g__Brevundimonas | D−Arabinose 5−phosphate | −0.197 | 0.271 | negtive |
| g__Gardnerella | D−Arabinose 5−phosphate | −0.196 | 0.275 | negtive |
| g__Leifsonia | D−Arabinose 5−phosphate | −0.142 | 0.432 | negtive |
| g__Vibrio | D−Arabinose 5−phosphate | −0.115 | 0.523 | negtive |
| g__Aeromonas | D−Arabinose 5−phosphate | −0.066 | 0.717 | negtive |
| g__Pseudoalteromonas | D−Arabinose 5−phosphate | −0.039 | 0.831 | negtive |
| g__Lactobacillus | D−Cysteine | −0.390 | 0.025 | negtive |
| g__Gardnerella | D−Cysteine | −0.389 | 0.025 | negtive |
| g__Mycoplasma | D−Cysteine | 0.351 | 0.045 | positive |
| g__Acinetobacter | D−Cysteine | −0.307 | 0.082 | negtive |
| g__Hydrogenophaga | D−Cysteine | −0.286 | 0.106 | negtive |
| g__Haemophilus | D−Cysteine | 0.268 | 0.132 | positive |
| g__Tropheryma | D−Cysteine | 0.248 | 0.163 | positive |
| g__Prevotella | D−Cysteine | 0.228 | 0.203 | positive |
| g__Moraxella | D−Cysteine | −0.194 | 0.280 | negtive |
| g__Porphyromonas | D−Cysteine | 0.149 | 0.406 | positive |
| g__Streptococcus | D−Cysteine | 0.144 | 0.423 | positive |
| g__Veillonella | D−Cysteine | 0.135 | 0.454 | positive |
| g__Leifsonia | D−Cysteine | −0.121 | 0.504 | negtive |
| g__Brevundimonas | D−Cysteine | −0.106 | 0.555 | negtive |
| g__Alloprevotella | D−Cysteine | 0.101 | 0.575 | positive |
| g__Pseudoalteromonas | D−Cysteine | 0.063 | 0.728 | positive |
| g__Neisseria | D−Cysteine | 0.055 | 0.760 | positive |
| g__Vibrio | D−Cysteine | −0.046 | 0.799 | negtive |
| g__Aeromonas | D−Cysteine | 0.032 | 0.860 | positive |
| g__Prevotella_7 | D−Cysteine | −0.013 | 0.943 | negtive |
| g__Hydrogenophaga | Formononetin | −0.611 | 0.000 | negtive |
| g__Acinetobacter | Formononetin | −0.593 | 0.000 | negtive |
| g__Mycoplasma | Formononetin | 0.419 | 0.015 | positive |
| g__Porphyromonas | Formononetin | 0.313 | 0.076 | positive |
| g__Haemophilus | Formononetin | 0.309 | 0.081 | positive |
| g__Gardnerella | Formononetin | −0.273 | 0.124 | negtive |
| g__Moraxella | Formononetin | −0.269 | 0.130 | negtive |
| g__Lactobacillus | Formononetin | −0.246 | 0.167 | negtive |
| g__Brevundimonas | Formononetin | −0.232 | 0.193 | negtive |
| g__Prevotella | Formononetin | 0.214 | 0.233 | positive |
| g__Veillonella | Formononetin | 0.207 | 0.248 | positive |
| g__Tropheryma | Formononetin | 0.185 | 0.304 | positive |
| g__Streptococcus | Formononetin | 0.164 | 0.360 | positive |
| g__Leifsonia | Formononetin | −0.149 | 0.407 | negtive |
| g__Neisseria | Formononetin | 0.073 | 0.686 | positive |
| g__Vibrio | Formononetin | −0.023 | 0.901 | negtive |
| g__Aeromonas | Formononetin | −0.022 | 0.905 | negtive |
| g__Prevotella_7 | Formononetin | 0.004 | 0.982 | positive |
| g__Alloprevotella | Formononetin | 0.003 | 0.985 | positive |
| g__Pseudoalteromonas | Formononetin | −0.002 | 0.993 | negtive |
| g__Hydrogenophaga | Glucose 6−phosphate | −0.741 | 0.000 | negtive |
| g__Acinetobacter | Glucose 6−phosphate | −0.650 | 0.000 | negtive |
| g__Porphyromonas | Glucose 6−phosphate | 0.452 | 0.008 | positive |
| g__Haemophilus | Glucose 6−phosphate | 0.443 | 0.010 | positive |
| g__Brevundimonas | Glucose 6−phosphate | −0.414 | 0.017 | negtive |
| g__Leifsonia | Glucose 6−phosphate | −0.361 | 0.039 | negtive |
| g__Moraxella | Glucose 6−phosphate | −0.294 | 0.097 | negtive |
| g__Veillonella | Glucose 6−phosphate | 0.289 | 0.103 | positive |
| g__Vibrio | Glucose 6−phosphate | −0.247 | 0.166 | negtive |
| g__Mycoplasma | Glucose 6−phosphate | 0.218 | 0.222 | positive |
| g__Aeromonas | Glucose 6−phosphate | −0.217 | 0.224 | negtive |
| g__Pseudoalteromonas | Glucose 6−phosphate | −0.208 | 0.245 | negtive |
| g__Tropheryma | Glucose 6−phosphate | 0.200 | 0.265 | positive |
| g__Gardnerella | Glucose 6−phosphate | −0.192 | 0.286 | negtive |
| g__Streptococcus | Glucose 6−phosphate | 0.182 | 0.310 | positive |
| g__Prevotella | Glucose 6−phosphate | 0.173 | 0.334 | positive |
| g__Neisseria | Glucose 6−phosphate | 0.166 | 0.357 | positive |
| g__Lactobacillus | Glucose 6−phosphate | −0.118 | 0.513 | negtive |
| g__Alloprevotella | Glucose 6−phosphate | 0.107 | 0.555 | positive |
| g__Prevotella_7 | Glucose 6−phosphate | 0.073 | 0.687 | positive |
| g__Tropheryma | Glycidol | 0.452 | 0.008 | positive |
| g__Prevotella | Glycidol | 0.379 | 0.029 | positive |
| g__Gardnerella | Glycidol | −0.357 | 0.041 | negtive |
| g__Prevotella_7 | Glycidol | 0.317 | 0.072 | positive |
| g__Haemophilus | Glycidol | 0.292 | 0.099 | positive |
| g__Veillonella | Glycidol | 0.284 | 0.109 | positive |
| g__Alloprevotella | Glycidol | 0.282 | 0.112 | positive |
| g__Lactobacillus | Glycidol | −0.277 | 0.119 | negtive |
| g__Neisseria | Glycidol | 0.266 | 0.134 | positive |
| g__Streptococcus | Glycidol | 0.229 | 0.200 | positive |
| g__Pseudoalteromonas | Glycidol | 0.212 | 0.237 | positive |
| g__Aeromonas | Glycidol | 0.186 | 0.299 | positive |
| g__Leifsonia | Glycidol | 0.163 | 0.364 | positive |
| g__Vibrio | Glycidol | 0.147 | 0.415 | positive |
| g__Brevundimonas | Glycidol | 0.132 | 0.461 | positive |
| g__Moraxella | Glycidol | 0.113 | 0.533 | positive |
| g__Porphyromonas | Glycidol | 0.110 | 0.544 | positive |
| g__Hydrogenophaga | Glycidol | 0.081 | 0.652 | positive |
| g__Mycoplasma | Glycidol | 0.074 | 0.683 | positive |
| g__Acinetobacter | Glycidol | 0.054 | 0.764 | positive |
| g__Mycoplasma | Glycine | 0.375 | 0.031 | positive |
| g__Tropheryma | Glycine | 0.281 | 0.114 | positive |
| g__Lactobacillus | Glycine | −0.265 | 0.135 | negtive |
| g__Hydrogenophaga | Glycine | −0.235 | 0.187 | negtive |
| g__Gardnerella | Glycine | −0.220 | 0.219 | negtive |
| g__Haemophilus | Glycine | 0.204 | 0.255 | positive |
| g__Vibrio | Glycine | −0.197 | 0.272 | negtive |
| g__Acinetobacter | Glycine | −0.193 | 0.282 | negtive |
| g__Neisseria | Glycine | 0.182 | 0.309 | positive |
| g__Alloprevotella | Glycine | 0.175 | 0.330 | positive |
| g__Porphyromonas | Glycine | 0.166 | 0.355 | positive |
| g__Prevotella | Glycine | 0.164 | 0.361 | positive |
| g__Brevundimonas | Glycine | −0.140 | 0.434 | negtive |
| g__Veillonella | Glycine | 0.140 | 0.436 | positive |
| g__Moraxella | Glycine | −0.119 | 0.508 | negtive |
| g__Aeromonas | Glycine | −0.111 | 0.539 | negtive |
| g__Leifsonia | Glycine | −0.107 | 0.553 | negtive |
| g__Streptococcus | Glycine | 0.100 | 0.581 | positive |
| g__Pseudoalteromonas | Glycine | −0.081 | 0.656 | negtive |
| g__Prevotella_7 | Glycine | 0.063 | 0.728 | positive |
| g__Mycoplasma | Glycolic acid | 0.427 | 0.013 | positive |
| g__Lactobacillus | Glycolic acid | −0.365 | 0.038 | negtive |
| g__Gardnerella | Glycolic acid | −0.364 | 0.037 | negtive |
| g__Tropheryma | Glycolic acid | 0.305 | 0.084 | positive |
| g__Hydrogenophaga | Glycolic acid | −0.240 | 0.178 | negtive |
| g__Haemophilus | Glycolic acid | 0.203 | 0.257 | positive |
| g__Neisseria | Glycolic acid | 0.179 | 0.320 | positive |
| g__Acinetobacter | Glycolic acid | −0.161 | 0.369 | negtive |
| g__Alloprevotella | Glycolic acid | 0.156 | 0.385 | positive |
| g__Veillonella | Glycolic acid | 0.144 | 0.426 | positive |
| g__Porphyromonas | Glycolic acid | 0.137 | 0.449 | positive |
| g__Prevotella | Glycolic acid | 0.134 | 0.456 | positive |
| g__Moraxella | Glycolic acid | −0.106 | 0.557 | negtive |
| g__Pseudoalteromonas | Glycolic acid | 0.077 | 0.672 | positive |
| g__Streptococcus | Glycolic acid | 0.067 | 0.712 | positive |
| g__Aeromonas | Glycolic acid | 0.037 | 0.836 | positive |
| g__Vibrio | Glycolic acid | −0.037 | 0.836 | negtive |
| g__Leifsonia | Glycolic acid | 0.007 | 0.971 | positive |
| g__Prevotella_7 | Glycolic acid | 0.002 | 0.991 | positive |
| g__Brevundimonas | Glycolic acid | −0.002 | 0.992 | negtive |
| g__Hydrogenophaga | Guanosine monophosphate | −0.502 | 0.003 | negtive |
| g__Brevundimonas | Guanosine monophosphate | −0.324 | 0.066 | negtive |
| g__Porphyromonas | Guanosine monophosphate | 0.309 | 0.080 | positive |
| g__Acinetobacter | Guanosine monophosphate | −0.301 | 0.089 | negtive |
| g__Haemophilus | Guanosine monophosphate | 0.289 | 0.103 | positive |
| g__Leifsonia | Guanosine monophosphate | −0.278 | 0.117 | negtive |
| g__Mycoplasma | Guanosine monophosphate | 0.255 | 0.152 | positive |
| g__Prevotella | Guanosine monophosphate | 0.150 | 0.406 | positive |
| g__Gardnerella | Guanosine monophosphate | −0.149 | 0.408 | negtive |
| g__Moraxella | Guanosine monophosphate | −0.134 | 0.458 | negtive |
| g__Vibrio | Guanosine monophosphate | −0.108 | 0.549 | negtive |
| g__Veillonella | Guanosine monophosphate | 0.098 | 0.586 | positive |
| g__Streptococcus | Guanosine monophosphate | 0.088 | 0.625 | positive |
| g__Prevotella_7 | Guanosine monophosphate | 0.066 | 0.714 | positive |
| g__Neisseria | Guanosine monophosphate | 0.065 | 0.721 | positive |
| g__Aeromonas | Guanosine monophosphate | −0.047 | 0.796 | negtive |
| g__Pseudoalteromonas | Guanosine monophosphate | −0.045 | 0.802 | negtive |
| g__Alloprevotella | Guanosine monophosphate | −0.040 | 0.825 | negtive |
| g__Tropheryma | Guanosine monophosphate | 0.027 | 0.881 | positive |
| g__Lactobacillus | Guanosine monophosphate | −0.026 | 0.887 | negtive |
| g__Hydrogenophaga | Hippuric acid | −0.386 | 0.027 | negtive |
| g__Leifsonia | Hippuric acid | −0.370 | 0.034 | negtive |
| g__Acinetobacter | Hippuric acid | −0.352 | 0.045 | negtive |
| g__Brevundimonas | Hippuric acid | −0.344 | 0.051 | negtive |
| g__Vibrio | Hippuric acid | −0.304 | 0.086 | negtive |
| g__Pseudoalteromonas | Hippuric acid | −0.299 | 0.092 | negtive |
| g__Aeromonas | Hippuric acid | −0.292 | 0.099 | negtive |
| g__Prevotella_7 | Hippuric acid | −0.266 | 0.135 | negtive |
| g__Alloprevotella | Hippuric acid | −0.188 | 0.294 | negtive |
| g__Veillonella | Hippuric acid | −0.183 | 0.308 | negtive |
| g__Streptococcus | Hippuric acid | −0.182 | 0.311 | negtive |
| g__Gardnerella | Hippuric acid | −0.147 | 0.415 | negtive |
| g__Moraxella | Hippuric acid | −0.131 | 0.469 | negtive |
| g__Neisseria | Hippuric acid | −0.117 | 0.517 | negtive |
| g__Mycoplasma | Hippuric acid | 0.110 | 0.543 | positive |
| g__Haemophilus | Hippuric acid | 0.083 | 0.647 | positive |
| g__Lactobacillus | Hippuric acid | 0.081 | 0.653 | positive |
| g__Porphyromonas | Hippuric acid | 0.073 | 0.687 | positive |
| g__Tropheryma | Hippuric acid | −0.068 | 0.705 | negtive |
| g__Prevotella | Hippuric acid | −0.033 | 0.855 | negtive |
| g__Porphyromonas | Imazamox | 0.568 | 0.001 | positive |
| g__Haemophilus | Imazamox | 0.558 | 0.001 | positive |
| g__Hydrogenophaga | Imazamox | −0.430 | 0.013 | negtive |
| g__Veillonella | Imazamox | 0.415 | 0.016 | positive |
| g__Prevotella_7 | Imazamox | 0.413 | 0.017 | positive |
| g__Acinetobacter | Imazamox | −0.374 | 0.033 | negtive |
| g__Neisseria | Imazamox | 0.362 | 0.038 | positive |
| g__Alloprevotella | Imazamox | 0.311 | 0.078 | positive |
| g__Prevotella | Imazamox | 0.299 | 0.091 | positive |
| g__Streptococcus | Imazamox | 0.296 | 0.094 | positive |
| g__Mycoplasma | Imazamox | 0.287 | 0.106 | positive |
| g__Lactobacillus | Imazamox | −0.269 | 0.130 | negtive |
| g__Gardnerella | Imazamox | −0.256 | 0.151 | negtive |
| g__Tropheryma | Imazamox | 0.227 | 0.205 | positive |
| g__Leifsonia | Imazamox | −0.075 | 0.677 | negtive |
| g__Brevundimonas | Imazamox | −0.058 | 0.746 | negtive |
| g__Moraxella | Imazamox | −0.020 | 0.914 | negtive |
| g__Vibrio | Imazamox | 0.019 | 0.917 | positive |
| g__Aeromonas | Imazamox | −0.016 | 0.931 | negtive |
| g__Pseudoalteromonas | Imazamox | 0.007 | 0.968 | positive |
| g__Haemophilus | Lenticin | 0.442 | 0.010 | positive |
| g__Porphyromonas | Lenticin | 0.336 | 0.056 | positive |
| g__Prevotella | Lenticin | 0.327 | 0.063 | positive |
| g__Acinetobacter | Lenticin | −0.314 | 0.075 | negtive |
| g__Brevundimonas | Lenticin | −0.252 | 0.157 | negtive |
| g__Leifsonia | Lenticin | −0.249 | 0.162 | negtive |
| g__Hydrogenophaga | Lenticin | −0.235 | 0.187 | negtive |
| g__Moraxella | Lenticin | −0.205 | 0.253 | negtive |
| g__Alloprevotella | Lenticin | 0.194 | 0.279 | positive |
| g__Neisseria | Lenticin | 0.190 | 0.289 | positive |
| g__Veillonella | Lenticin | 0.188 | 0.295 | positive |
| g__Gardnerella | Lenticin | −0.126 | 0.484 | negtive |
| g__Mycoplasma | Lenticin | 0.118 | 0.515 | positive |
| g__Aeromonas | Lenticin | −0.113 | 0.530 | negtive |
| g__Streptococcus | Lenticin | 0.112 | 0.534 | positive |
| g__Prevotella_7 | Lenticin | 0.087 | 0.630 | positive |
| g__Vibrio | Lenticin | −0.085 | 0.640 | negtive |
| g__Pseudoalteromonas | Lenticin | −0.082 | 0.652 | negtive |
| g__Lactobacillus | Lenticin | 0.032 | 0.861 | positive |
| g__Tropheryma | Lenticin | 0.021 | 0.909 | positive |
| g__Acinetobacter | Malondialdehyde | −0.621 | 0.000 | negtive |
| g__Hydrogenophaga | Malondialdehyde | −0.589 | 0.000 | negtive |
| g__Porphyromonas | Malondialdehyde | 0.435 | 0.011 | positive |
| g__Veillonella | Malondialdehyde | 0.386 | 0.027 | positive |
| g__Vibrio | Malondialdehyde | −0.359 | 0.040 | negtive |
| g__Brevundimonas | Malondialdehyde | −0.354 | 0.044 | negtive |
| g__Aeromonas | Malondialdehyde | −0.317 | 0.072 | negtive |
| g__Leifsonia | Malondialdehyde | −0.303 | 0.087 | negtive |
| g__Pseudoalteromonas | Malondialdehyde | −0.297 | 0.093 | negtive |
| g__Moraxella | Malondialdehyde | −0.274 | 0.122 | negtive |
| g__Haemophilus | Malondialdehyde | 0.274 | 0.122 | positive |
| g__Streptococcus | Malondialdehyde | 0.271 | 0.128 | positive |
| g__Mycoplasma | Malondialdehyde | 0.256 | 0.150 | positive |
| g__Lactobacillus | Malondialdehyde | −0.252 | 0.156 | negtive |
| g__Alloprevotella | Malondialdehyde | 0.181 | 0.313 | positive |
| g__Prevotella | Malondialdehyde | 0.147 | 0.415 | positive |
| g__Gardnerella | Malondialdehyde | −0.139 | 0.440 | negtive |
| g__Tropheryma | Malondialdehyde | 0.110 | 0.541 | positive |
| g__Neisseria | Malondialdehyde | 0.086 | 0.634 | positive |
| g__Prevotella_7 | Malondialdehyde | 0.051 | 0.779 | positive |
| g__Hydrogenophaga | MANNOSE | −0.534 | 0.002 | negtive |
| g__Acinetobacter | MANNOSE | −0.533 | 0.002 | negtive |
| g__Porphyromonas | MANNOSE | 0.510 | 0.002 | positive |
| g__Veillonella | MANNOSE | 0.462 | 0.007 | positive |
| g__Haemophilus | MANNOSE | 0.357 | 0.041 | positive |
| g__Lactobacillus | MANNOSE | −0.349 | 0.047 | negtive |
| g__Mycoplasma | MANNOSE | 0.313 | 0.076 | positive |
| g__Streptococcus | MANNOSE | 0.295 | 0.095 | positive |
| g__Prevotella | MANNOSE | 0.289 | 0.103 | positive |
| g__Alloprevotella | MANNOSE | 0.283 | 0.110 | positive |
| g__Vibrio | MANNOSE | −0.278 | 0.117 | negtive |
| g__Brevundimonas | MANNOSE | −0.274 | 0.123 | negtive |
| g__Leifsonia | MANNOSE | −0.229 | 0.200 | negtive |
| g__Aeromonas | MANNOSE | −0.229 | 0.200 | negtive |
| g__Neisseria | MANNOSE | 0.227 | 0.205 | positive |
| g__Pseudoalteromonas | MANNOSE | −0.203 | 0.256 | negtive |
| g__Moraxella | MANNOSE | −0.179 | 0.318 | negtive |
| g__Prevotella_7 | MANNOSE | 0.142 | 0.429 | positive |
| g__Tropheryma | MANNOSE | 0.137 | 0.449 | positive |
| g__Gardnerella | MANNOSE | −0.132 | 0.464 | negtive |
| g__Moraxella | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.403 | 0.020 | negtive |
| g__Acinetobacter | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.363 | 0.039 | negtive |
| g__Hydrogenophaga | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.275 | 0.121 | negtive |
| g__Lactobacillus | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.243 | 0.173 | negtive |
| g__Brevundimonas | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.237 | 0.184 | negtive |
| g__Leifsonia | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.223 | 0.212 | negtive |
| g__Mycoplasma | N−Acetyl−alpha−D−glucosamine 1−phosphate | 0.206 | 0.250 | positive |
| g__Aeromonas | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.188 | 0.295 | negtive |
| g__Tropheryma | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.185 | 0.301 | negtive |
| g__Gardnerella | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.172 | 0.340 | negtive |
| g__Vibrio | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.164 | 0.363 | negtive |
| g__Pseudoalteromonas | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.149 | 0.408 | negtive |
| g__Porphyromonas | N−Acetyl−alpha−D−glucosamine 1−phosphate | 0.134 | 0.457 | positive |
| g__Haemophilus | N−Acetyl−alpha−D−glucosamine 1−phosphate | 0.134 | 0.457 | positive |
| g__Neisseria | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.109 | 0.547 | negtive |
| g__Prevotella_7 | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.096 | 0.596 | negtive |
| g__Alloprevotella | N−Acetyl−alpha−D−glucosamine 1−phosphate | 0.072 | 0.690 | positive |
| g__Veillonella | N−Acetyl−alpha−D−glucosamine 1−phosphate | 0.065 | 0.719 | positive |
| g__Streptococcus | N−Acetyl−alpha−D−glucosamine 1−phosphate | −0.050 | 0.784 | negtive |
| g__Prevotella | N−Acetyl−alpha−D−glucosamine 1−phosphate | 0.015 | 0.935 | positive |
| g__Acinetobacter | N−Acetyl−D−tryptophan | −0.364 | 0.038 | negtive |
| g__Vibrio | N−Acetyl−D−tryptophan | −0.329 | 0.062 | negtive |
| g__Hydrogenophaga | N−Acetyl−D−tryptophan | −0.327 | 0.064 | negtive |
| g__Brevundimonas | N−Acetyl−D−tryptophan | −0.318 | 0.072 | negtive |
| g__Pseudoalteromonas | N−Acetyl−D−tryptophan | −0.305 | 0.084 | negtive |
| g__Aeromonas | N−Acetyl−D−tryptophan | −0.300 | 0.090 | negtive |
| g__Leifsonia | N−Acetyl−D−tryptophan | −0.221 | 0.217 | negtive |
| g__Moraxella | N−Acetyl−D−tryptophan | −0.180 | 0.316 | negtive |
| g__Tropheryma | N−Acetyl−D−tryptophan | 0.153 | 0.397 | positive |
| g__Lactobacillus | N−Acetyl−D−tryptophan | 0.151 | 0.401 | positive |
| g__Gardnerella | N−Acetyl−D−tryptophan | −0.133 | 0.462 | negtive |
| g__Haemophilus | N−Acetyl−D−tryptophan | 0.109 | 0.545 | positive |
| g__Neisseria | N−Acetyl−D−tryptophan | −0.075 | 0.680 | negtive |
| g__Alloprevotella | N−Acetyl−D−tryptophan | −0.066 | 0.714 | negtive |
| g__Prevotella_7 | N−Acetyl−D−tryptophan | −0.051 | 0.777 | negtive |
| g__Veillonella | N−Acetyl−D−tryptophan | 0.036 | 0.843 | positive |
| g__Streptococcus | N−Acetyl−D−tryptophan | 0.025 | 0.889 | positive |
| g__Porphyromonas | N−Acetyl−D−tryptophan | −0.014 | 0.940 | negtive |
| g__Prevotella | N−Acetyl−D−tryptophan | −0.001 | 0.994 | negtive |
| g__Mycoplasma | N−Acetyl−D−tryptophan | −0.001 | 0.997 | negtive |
| g__Mycoplasma | N−Acetyl−L−cysteine | 0.379 | 0.030 | positive |
| g__Hydrogenophaga | N−Acetyl−L−cysteine | −0.332 | 0.060 | negtive |
| g__Prevotella | N−Acetyl−L−cysteine | 0.328 | 0.062 | positive |
| g__Porphyromonas | N−Acetyl−L−cysteine | 0.285 | 0.107 | positive |
| g__Acinetobacter | N−Acetyl−L−cysteine | −0.280 | 0.114 | negtive |
| g__Haemophilus | N−Acetyl−L−cysteine | 0.260 | 0.144 | positive |
| g__Lactobacillus | N−Acetyl−L−cysteine | −0.218 | 0.222 | negtive |
| g__Veillonella | N−Acetyl−L−cysteine | 0.180 | 0.317 | positive |
| g__Gardnerella | N−Acetyl−L−cysteine | −0.166 | 0.355 | negtive |
| g__Prevotella_7 | N−Acetyl−L−cysteine | 0.121 | 0.504 | positive |
| g__Neisseria | N−Acetyl−L−cysteine | 0.120 | 0.508 | positive |
| g__Alloprevotella | N−Acetyl−L−cysteine | 0.112 | 0.536 | positive |
| g__Tropheryma | N−Acetyl−L−cysteine | 0.102 | 0.573 | positive |
| g__Leifsonia | N−Acetyl−L−cysteine | −0.095 | 0.600 | negtive |
| g__Moraxella | N−Acetyl−L−cysteine | −0.089 | 0.621 | negtive |
| g__Brevundimonas | N−Acetyl−L−cysteine | −0.073 | 0.685 | negtive |
| g__Pseudoalteromonas | N−Acetyl−L−cysteine | 0.058 | 0.746 | positive |
| g__Aeromonas | N−Acetyl−L−cysteine | −0.008 | 0.965 | negtive |
| g__Streptococcus | N−Acetyl−L−cysteine | −0.003 | 0.987 | negtive |
| g__Vibrio | N−Acetyl−L−cysteine | −0.001 | 0.996 | negtive |
| g__Streptococcus | N−Acetyl−L−phenylalanine | 0.463 | 0.007 | positive |
| g__Porphyromonas | N−Acetyl−L−phenylalanine | 0.424 | 0.014 | positive |
| g__Lactobacillus | N−Acetyl−L−phenylalanine | −0.404 | 0.020 | negtive |
| g__Veillonella | N−Acetyl−L−phenylalanine | 0.391 | 0.025 | positive |
| g__Mycoplasma | N−Acetyl−L−phenylalanine | 0.325 | 0.065 | positive |
| g__Acinetobacter | N−Acetyl−L−phenylalanine | −0.319 | 0.071 | negtive |
| g__Alloprevotella | N−Acetyl−L−phenylalanine | 0.301 | 0.089 | positive |
| g__Haemophilus | N−Acetyl−L−phenylalanine | 0.281 | 0.113 | positive |
| g__Neisseria | N−Acetyl−L−phenylalanine | 0.281 | 0.113 | positive |
| g__Prevotella | N−Acetyl−L−phenylalanine | 0.257 | 0.149 | positive |
| g__Tropheryma | N−Acetyl−L−phenylalanine | 0.237 | 0.185 | positive |
| g__Hydrogenophaga | N−Acetyl−L−phenylalanine | −0.233 | 0.191 | negtive |
| g__Gardnerella | N−Acetyl−L−phenylalanine | −0.212 | 0.237 | negtive |
| g__Prevotella_7 | N−Acetyl−L−phenylalanine | 0.153 | 0.397 | positive |
| g__Vibrio | N−Acetyl−L−phenylalanine | −0.120 | 0.506 | negtive |
| g__Moraxella | N−Acetyl−L−phenylalanine | 0.044 | 0.807 | positive |
| g__Pseudoalteromonas | N−Acetyl−L−phenylalanine | −0.036 | 0.843 | negtive |
| g__Aeromonas | N−Acetyl−L−phenylalanine | −0.022 | 0.901 | negtive |
| g__Brevundimonas | N−Acetyl−L−phenylalanine | −0.020 | 0.911 | negtive |
| g__Leifsonia | N−Acetyl−L−phenylalanine | −0.012 | 0.948 | negtive |
| g__Haemophilus | Neobyakangelicol | 0.521 | 0.002 | positive |
| g__Porphyromonas | Neobyakangelicol | 0.354 | 0.043 | positive |
| g__Leifsonia | Neobyakangelicol | −0.310 | 0.079 | negtive |
| g__Gardnerella | Neobyakangelicol | −0.310 | 0.079 | negtive |
| g__Acinetobacter | Neobyakangelicol | −0.274 | 0.122 | negtive |
| g__Veillonella | Neobyakangelicol | 0.263 | 0.139 | positive |
| g__Streptococcus | Neobyakangelicol | 0.250 | 0.161 | positive |
| g__Hydrogenophaga | Neobyakangelicol | −0.248 | 0.164 | negtive |
| g__Tropheryma | Neobyakangelicol | 0.223 | 0.213 | positive |
| g__Brevundimonas | Neobyakangelicol | −0.221 | 0.216 | negtive |
| g__Prevotella | Neobyakangelicol | 0.190 | 0.290 | positive |
| g__Prevotella_7 | Neobyakangelicol | 0.187 | 0.299 | positive |
| g__Lactobacillus | Neobyakangelicol | −0.175 | 0.328 | negtive |
| g__Vibrio | Neobyakangelicol | −0.146 | 0.418 | negtive |
| g__Pseudoalteromonas | Neobyakangelicol | −0.111 | 0.537 | negtive |
| g__Alloprevotella | Neobyakangelicol | 0.106 | 0.557 | positive |
| g__Aeromonas | Neobyakangelicol | −0.082 | 0.649 | negtive |
| g__Neisseria | Neobyakangelicol | 0.081 | 0.654 | positive |
| g__Moraxella | Neobyakangelicol | −0.057 | 0.754 | negtive |
| g__Mycoplasma | Neobyakangelicol | −0.022 | 0.903 | negtive |
| g__Mycoplasma | N−Formylmethionine | 0.451 | 0.008 | positive |
| g__Gardnerella | N−Formylmethionine | −0.394 | 0.023 | negtive |
| g__Acinetobacter | N−Formylmethionine | −0.369 | 0.035 | negtive |
| g__Lactobacillus | N−Formylmethionine | −0.363 | 0.038 | negtive |
| g__Hydrogenophaga | N−Formylmethionine | −0.358 | 0.042 | negtive |
| g__Haemophilus | N−Formylmethionine | 0.276 | 0.121 | positive |
| g__Tropheryma | N−Formylmethionine | 0.214 | 0.231 | positive |
| g__Moraxella | N−Formylmethionine | −0.203 | 0.257 | negtive |
| g__Porphyromonas | N−Formylmethionine | 0.202 | 0.261 | positive |
| g__Streptococcus | N−Formylmethionine | 0.165 | 0.358 | positive |
| g__Neisseria | N−Formylmethionine | 0.117 | 0.517 | positive |
| g__Alloprevotella | N−Formylmethionine | 0.115 | 0.523 | positive |
| g__Veillonella | N−Formylmethionine | 0.108 | 0.549 | positive |
| g__Pseudoalteromonas | N−Formylmethionine | 0.105 | 0.561 | positive |
| g__Aeromonas | N−Formylmethionine | 0.103 | 0.569 | positive |
| g__Prevotella | N−Formylmethionine | 0.084 | 0.642 | positive |
| g__Vibrio | N−Formylmethionine | 0.046 | 0.801 | positive |
| g__Prevotella_7 | N−Formylmethionine | −0.037 | 0.840 | negtive |
| g__Brevundimonas | N−Formylmethionine | −0.030 | 0.868 | negtive |
| g__Leifsonia | N−Formylmethionine | −0.028 | 0.877 | negtive |
| g__Acinetobacter | N−Nitrosodimethylamine | 0.399 | 0.022 | positive |
| g__Hydrogenophaga | N−Nitrosodimethylamine | 0.378 | 0.031 | positive |
| g__Haemophilus | N−Nitrosodimethylamine | −0.280 | 0.114 | negtive |
| g__Leifsonia | N−Nitrosodimethylamine | 0.213 | 0.233 | positive |
| g__Mycoplasma | N−Nitrosodimethylamine | −0.197 | 0.272 | negtive |
| g__Brevundimonas | N−Nitrosodimethylamine | 0.179 | 0.318 | positive |
| g__Moraxella | N−Nitrosodimethylamine | 0.159 | 0.377 | positive |
| g__Prevotella_7 | N−Nitrosodimethylamine | 0.149 | 0.409 | positive |
| g__Veillonella | N−Nitrosodimethylamine | 0.139 | 0.442 | positive |
| g__Streptococcus | N−Nitrosodimethylamine | 0.112 | 0.536 | positive |
| g__Porphyromonas | N−Nitrosodimethylamine | −0.110 | 0.541 | negtive |
| g__Gardnerella | N−Nitrosodimethylamine | −0.077 | 0.669 | negtive |
| g__Aeromonas | N−Nitrosodimethylamine | 0.056 | 0.758 | positive |
| g__Lactobacillus | N−Nitrosodimethylamine | 0.052 | 0.771 | positive |
| g__Alloprevotella | N−Nitrosodimethylamine | −0.050 | 0.780 | negtive |
| g__Pseudoalteromonas | N−Nitrosodimethylamine | 0.025 | 0.892 | positive |
| g__Vibrio | N−Nitrosodimethylamine | 0.021 | 0.908 | positive |
| g__Tropheryma | N−Nitrosodimethylamine | 0.020 | 0.910 | positive |
| g__Prevotella | N−Nitrosodimethylamine | −0.015 | 0.934 | negtive |
| g__Neisseria | N−Nitrosodimethylamine | −0.009 | 0.960 | negtive |
| g__Lactobacillus | O−Phospho−L−serine | −0.261 | 0.142 | negtive |
| g__Acinetobacter | O−Phospho−L−serine | −0.259 | 0.145 | negtive |
| g__Hydrogenophaga | O−Phospho−L−serine | −0.247 | 0.165 | negtive |
| g__Mycoplasma | O−Phospho−L−serine | 0.216 | 0.228 | positive |
| g__Tropheryma | O−Phospho−L−serine | 0.207 | 0.249 | positive |
| g__Streptococcus | O−Phospho−L−serine | −0.192 | 0.286 | negtive |
| g__Prevotella_7 | O−Phospho−L−serine | −0.142 | 0.430 | negtive |
| g__Moraxella | O−Phospho−L−serine | −0.119 | 0.511 | negtive |
| g__Neisseria | O−Phospho−L−serine | −0.090 | 0.620 | negtive |
| g__Gardnerella | O−Phospho−L−serine | −0.090 | 0.620 | negtive |
| g__Vibrio | O−Phospho−L−serine | −0.078 | 0.665 | negtive |
| g__Aeromonas | O−Phospho−L−serine | −0.058 | 0.748 | negtive |
| g__Prevotella | O−Phospho−L−serine | 0.052 | 0.774 | positive |
| g__Leifsonia | O−Phospho−L−serine | −0.052 | 0.774 | negtive |
| g__Brevundimonas | O−Phospho−L−serine | −0.048 | 0.788 | negtive |
| g__Alloprevotella | O−Phospho−L−serine | −0.034 | 0.850 | negtive |
| g__Porphyromonas | O−Phospho−L−serine | −0.033 | 0.854 | negtive |
| g__Veillonella | O−Phospho−L−serine | 0.012 | 0.948 | positive |
| g__Pseudoalteromonas | O−Phospho−L−serine | −0.002 | 0.993 | negtive |
| g__Haemophilus | O−Phospho−L−serine | −0.001 | 0.995 | negtive |
| g__Hydrogenophaga | Pelargonidin | −0.542 | 0.001 | negtive |
| g__Acinetobacter | Pelargonidin | −0.429 | 0.013 | negtive |
| g__Mycoplasma | Pelargonidin | 0.423 | 0.014 | positive |
| g__Moraxella | Pelargonidin | −0.417 | 0.016 | negtive |
| g__Porphyromonas | Pelargonidin | 0.335 | 0.057 | positive |
| g__Brevundimonas | Pelargonidin | −0.321 | 0.069 | negtive |
| g__Haemophilus | Pelargonidin | 0.217 | 0.224 | positive |
| g__Prevotella | Pelargonidin | 0.217 | 0.225 | positive |
| g__Gardnerella | Pelargonidin | −0.197 | 0.272 | negtive |
| g__Veillonella | Pelargonidin | 0.162 | 0.368 | positive |
| g__Leifsonia | Pelargonidin | −0.151 | 0.403 | negtive |
| g__Alloprevotella | Pelargonidin | 0.143 | 0.427 | positive |
| g__Aeromonas | Pelargonidin | −0.126 | 0.483 | negtive |
| g__Lactobacillus | Pelargonidin | −0.119 | 0.508 | negtive |
| g__Vibrio | Pelargonidin | −0.094 | 0.602 | negtive |
| g__Neisseria | Pelargonidin | 0.082 | 0.651 | positive |
| g__Tropheryma | Pelargonidin | −0.067 | 0.711 | negtive |
| g__Pseudoalteromonas | Pelargonidin | −0.065 | 0.718 | negtive |
| g__Streptococcus | Pelargonidin | 0.006 | 0.973 | positive |
| g__Prevotella_7 | Pelargonidin | −0.001 | 0.996 | negtive |
| g__Hydrogenophaga | Pentedrone | 0.607 | 0.000 | positive |
| g__Acinetobacter | Pentedrone | 0.518 | 0.002 | positive |
| g__Moraxella | Pentedrone | 0.379 | 0.030 | positive |
| g__Brevundimonas | Pentedrone | 0.346 | 0.049 | positive |
| g__Haemophilus | Pentedrone | −0.328 | 0.062 | negtive |
| g__Porphyromonas | Pentedrone | −0.316 | 0.073 | negtive |
| g__Leifsonia | Pentedrone | 0.253 | 0.156 | positive |
| g__Mycoplasma | Pentedrone | −0.220 | 0.219 | negtive |
| g__Aeromonas | Pentedrone | 0.141 | 0.435 | positive |
| g__Tropheryma | Pentedrone | 0.135 | 0.455 | positive |
| g__Prevotella | Pentedrone | −0.122 | 0.500 | negtive |
| g__Pseudoalteromonas | Pentedrone | 0.116 | 0.520 | positive |
| g__Veillonella | Pentedrone | −0.115 | 0.526 | negtive |
| g__Gardnerella | Pentedrone | 0.108 | 0.548 | positive |
| g__Vibrio | Pentedrone | 0.104 | 0.566 | positive |
| g__Neisseria | Pentedrone | −0.084 | 0.643 | negtive |
| g__Alloprevotella | Pentedrone | −0.069 | 0.703 | negtive |
| g__Lactobacillus | Pentedrone | −0.045 | 0.804 | negtive |
| g__Streptococcus | Pentedrone | −0.029 | 0.872 | negtive |
| g__Prevotella_7 | Pentedrone | 0.012 | 0.946 | positive |
| g__Mycoplasma | PROPANIL | 0.304 | 0.086 | positive |
| g__Moraxella | PROPANIL | −0.252 | 0.157 | negtive |
| g__Acinetobacter | PROPANIL | −0.181 | 0.312 | negtive |
| g__Streptococcus | PROPANIL | −0.178 | 0.323 | negtive |
| g__Hydrogenophaga | PROPANIL | −0.168 | 0.349 | negtive |
| g__Prevotella | PROPANIL | 0.107 | 0.553 | positive |
| g__Leifsonia | PROPANIL | 0.058 | 0.748 | positive |
| g__Haemophilus | PROPANIL | 0.051 | 0.779 | positive |
| g__Gardnerella | PROPANIL | −0.050 | 0.784 | negtive |
| g__Lactobacillus | PROPANIL | −0.044 | 0.808 | negtive |
| g__Aeromonas | PROPANIL | −0.038 | 0.834 | negtive |
| g__Prevotella_7 | PROPANIL | 0.036 | 0.844 | positive |
| g__Vibrio | PROPANIL | 0.034 | 0.852 | positive |
| g__Brevundimonas | PROPANIL | −0.032 | 0.858 | negtive |
| g__Pseudoalteromonas | PROPANIL | 0.030 | 0.869 | positive |
| g__Alloprevotella | PROPANIL | 0.025 | 0.888 | positive |
| g__Porphyromonas | PROPANIL | 0.024 | 0.894 | positive |
| g__Veillonella | PROPANIL | 0.021 | 0.908 | positive |
| g__Tropheryma | PROPANIL | 0.003 | 0.985 | positive |
| g__Neisseria | PROPANIL | 0 | 1 | |
| g__Porphyromonas | Propyzamide | 0.543 | 0.001 | positive |
| g__Haemophilus | Propyzamide | 0.531 | 0.001 | positive |
| g__Hydrogenophaga | Propyzamide | −0.422 | 0.015 | negtive |
| g__Prevotella | Propyzamide | 0.421 | 0.015 | positive |
| g__Alloprevotella | Propyzamide | 0.386 | 0.027 | positive |
| g__Veillonella | Propyzamide | 0.369 | 0.034 | positive |
| g__Acinetobacter | Propyzamide | −0.362 | 0.039 | negtive |
| g__Neisseria | Propyzamide | 0.339 | 0.054 | positive |
| g__Moraxella | Propyzamide | −0.313 | 0.076 | negtive |
| g__Brevundimonas | Propyzamide | −0.287 | 0.105 | negtive |
| g__Prevotella_7 | Propyzamide | 0.261 | 0.142 | positive |
| g__Tropheryma | Propyzamide | −0.204 | 0.255 | negtive |
| g__Gardnerella | Propyzamide | −0.203 | 0.256 | negtive |
| g__Leifsonia | Propyzamide | −0.185 | 0.302 | negtive |
| g__Vibrio | Propyzamide | −0.152 | 0.400 | negtive |
| g__Streptococcus | Propyzamide | 0.141 | 0.434 | positive |
| g__Aeromonas | Propyzamide | −0.135 | 0.452 | negtive |
| g__Pseudoalteromonas | Propyzamide | −0.103 | 0.569 | negtive |
| g__Lactobacillus | Propyzamide | −0.058 | 0.746 | negtive |
| g__Mycoplasma | Propyzamide | 0.039 | 0.828 | positive |
| g__Mycoplasma | Pyroglutamic acid | 0.353 | 0.044 | positive |
| g__Lactobacillus | Pyroglutamic acid | −0.341 | 0.053 | negtive |
| g__Tropheryma | Pyroglutamic acid | 0.267 | 0.132 | positive |
| g__Hydrogenophaga | Pyroglutamic acid | −0.25 | 0.160 | negtive |
| g__Vibrio | Pyroglutamic acid | −0.236 | 0.186 | negtive |
| g__Acinetobacter | Pyroglutamic acid | −0.211 | 0.239 | negtive |
| g__Haemophilus | Pyroglutamic acid | 0.208 | 0.245 | positive |
| g__Alloprevotella | Pyroglutamic acid | 0.205 | 0.251 | positive |
| g__Gardnerella | Pyroglutamic acid | −0.203 | 0.256 | negtive |
| g__Porphyromonas | Pyroglutamic acid | 0.193 | 0.282 | positive |
| g__Prevotella | Pyroglutamic acid | 0.190 | 0.290 | positive |
| g__Neisseria | Pyroglutamic acid | 0.185 | 0.301 | positive |
| g__Veillonella | Pyroglutamic acid | 0.175 | 0.329 | positive |
| g__Streptococcus | Pyroglutamic acid | 0.149 | 0.407 | positive |
| g__Aeromonas | Pyroglutamic acid | −0.149 | 0.408 | negtive |
| g__Brevundimonas | Pyroglutamic acid | −0.108 | 0.547 | negtive |
| g__Pseudoalteromonas | Pyroglutamic acid | −0.106 | 0.557 | negtive |
| g__Leifsonia | Pyroglutamic acid | −0.098 | 0.589 | negtive |
| g__Moraxella | Pyroglutamic acid | −0.052 | 0.773 | negtive |
| g__Prevotella_7 | Pyroglutamic acid | 0.043 | 0.811 | positive |
| g__Hydrogenophaga | S−adenosylmethionine | −0.464 | 0.007 | negtive |
| g__Prevotella_7 | S−adenosylmethionine | −0.426 | 0.013 | negtive |
| g__Acinetobacter | S−adenosylmethionine | −0.390 | 0.026 | negtive |
| g__Leifsonia | S−adenosylmethionine | −0.387 | 0.026 | negtive |
| g__Brevundimonas | S−adenosylmethionine | −0.372 | 0.034 | negtive |
| g__Streptococcus | S−adenosylmethionine | −0.358 | 0.041 | negtive |
| g__Veillonella | S−adenosylmethionine | −0.339 | 0.054 | negtive |
| g__Lactobacillus | S−adenosylmethionine | 0.311 | 0.079 | positive |
| g__Alloprevotella | S−adenosylmethionine | −0.306 | 0.083 | negtive |
| g__Moraxella | S−adenosylmethionine | −0.301 | 0.088 | negtive |
| g__Vibrio | S−adenosylmethionine | −0.263 | 0.138 | negtive |
| g__Prevotella | S−adenosylmethionine | −0.204 | 0.256 | negtive |
| g__Neisseria | S−adenosylmethionine | −0.184 | 0.304 | negtive |
| g__Aeromonas | S−adenosylmethionine | −0.183 | 0.307 | negtive |
| g__Pseudoalteromonas | S−adenosylmethionine | −0.168 | 0.351 | negtive |
| g__Porphyromonas | S−adenosylmethionine | −0.120 | 0.505 | negtive |
| g__Haemophilus | S−adenosylmethionine | 0.069 | 0.705 | positive |
| g__Mycoplasma | S−adenosylmethionine | 0.067 | 0.709 | positive |
| g__Tropheryma | S−adenosylmethionine | −0.016 | 0.931 | negtive |
| g__Gardnerella | S−adenosylmethionine | 0.011 | 0.951 | positive |
| g__Hydrogenophaga | Sedoheptulose 7−phosphate | −0.699 | 0.000 | negtive |
| g__Acinetobacter | Sedoheptulose 7−phosphate | −0.541 | 0.001 | negtive |
| g__Brevundimonas | Sedoheptulose 7−phosphate | −0.514 | 0.003 | negtive |
| g__Leifsonia | Sedoheptulose 7−phosphate | −0.452 | 0.008 | negtive |
| g__Porphyromonas | Sedoheptulose 7−phosphate | 0.438 | 0.011 | positive |
| g__Haemophilus | Sedoheptulose 7−phosphate | 0.419 | 0.015 | positive |
| g__Vibrio | Sedoheptulose 7−phosphate | −0.408 | 0.018 | negtive |
| g__Aeromonas | Sedoheptulose 7−phosphate | −0.391 | 0.025 | negtive |
| g__Pseudoalteromonas | Sedoheptulose 7−phosphate | −0.381 | 0.029 | negtive |
| g__Moraxella | Sedoheptulose 7−phosphate | −0.303 | 0.087 | negtive |
| g__Veillonella | Sedoheptulose 7−phosphate | 0.195 | 0.276 | positive |
| g__Prevotella | Sedoheptulose 7−phosphate | 0.191 | 0.287 | positive |
| g__Neisseria | Sedoheptulose 7−phosphate | 0.137 | 0.446 | positive |
| g__Gardnerella | Sedoheptulose 7−phosphate | −0.130 | 0.470 | negtive |
| g__Streptococcus | Sedoheptulose 7−phosphate | 0.119 | 0.511 | positive |
| g__Alloprevotella | Sedoheptulose 7−phosphate | 0.093 | 0.605 | positive |
| g__Lactobacillus | Sedoheptulose 7−phosphate | 0.053 | 0.768 | positive |
| g__Mycoplasma | Sedoheptulose 7−phosphate | 0.052 | 0.772 | positive |
| g__Tropheryma | Sedoheptulose 7−phosphate | 0.023 | 0.897 | positive |
| g__Prevotella_7 | Sedoheptulose 7−phosphate | 0.017 | 0.924 | positive |
| g__Vibrio | Serotonin | 0.334 | 0.057 | positive |
| g__Leifsonia | Serotonin | 0.290 | 0.101 | positive |
| g__Pseudoalteromonas | Serotonin | 0.266 | 0.134 | positive |
| g__Aeromonas | Serotonin | 0.238 | 0.182 | positive |
| g__Neisseria | Serotonin | 0.229 | 0.201 | positive |
| g__Prevotella_7 | Serotonin | 0.185 | 0.302 | positive |
| g__Hydrogenophaga | Serotonin | 0.171 | 0.339 | positive |
| g__Brevundimonas | Serotonin | 0.160 | 0.373 | positive |
| g__Acinetobacter | Serotonin | 0.154 | 0.390 | positive |
| g__Alloprevotella | Serotonin | 0.153 | 0.394 | positive |
| g__Porphyromonas | Serotonin | 0.111 | 0.540 | positive |
| g__Gardnerella | Serotonin | 0.110 | 0.544 | positive |
| g__Prevotella | Serotonin | 0.094 | 0.603 | positive |
| g__Veillonella | Serotonin | 0.080 | 0.656 | positive |
| g__Mycoplasma | Serotonin | −0.069 | 0.703 | negtive |
| g__Haemophilus | Serotonin | 0.056 | 0.756 | positive |
| g__Streptococcus | Serotonin | 0.046 | 0.800 | positive |
| g__Tropheryma | Serotonin | −0.038 | 0.835 | negtive |
| g__Lactobacillus | Serotonin | 0.035 | 0.845 | positive |
| g__Moraxella | Serotonin | −0.007 | 0.971 | negtive |
| g__Mycoplasma | Sojagol | −0.423 | 0.014 | negtive |
| g__Acinetobacter | Sojagol | 0.366 | 0.037 | positive |
| g__Lactobacillus | Sojagol | 0.357 | 0.042 | positive |
| g__Hydrogenophaga | Sojagol | 0.317 | 0.073 | positive |
| g__Moraxella | Sojagol | 0.295 | 0.095 | positive |
| g__Porphyromonas | Sojagol | −0.252 | 0.158 | negtive |
| g__Gardnerella | Sojagol | 0.236 | 0.185 | positive |
| g__Veillonella | Sojagol | −0.232 | 0.193 | negtive |
| g__Tropheryma | Sojagol | −0.148 | 0.411 | negtive |
| g__Haemophilus | Sojagol | −0.141 | 0.434 | negtive |
| g__Brevundimonas | Sojagol | 0.101 | 0.576 | positive |
| g__Leifsonia | Sojagol | 0.094 | 0.602 | positive |
| g__Pseudoalteromonas | Sojagol | −0.078 | 0.668 | negtive |
| g__Streptococcus | Sojagol | −0.072 | 0.692 | negtive |
| g__Prevotella | Sojagol | −0.063 | 0.726 | negtive |
| g__Aeromonas | Sojagol | −0.063 | 0.727 | negtive |
| g__Vibrio | Sojagol | −0.063 | 0.730 | negtive |
| g__Alloprevotella | Sojagol | −0.039 | 0.828 | negtive |
| g__Prevotella_7 | Sojagol | −0.008 | 0.963 | negtive |
| g__Neisseria | Sojagol | −0.004 | 0.981 | negtive |
| g__Mycoplasma | Thioacetamide | 0.393 | 0.024 | positive |
| g__Hydrogenophaga | Thioacetamide | −0.308 | 0.082 | negtive |
| g__Acinetobacter | Thioacetamide | −0.264 | 0.137 | negtive |
| g__Vibrio | Thioacetamide | −0.259 | 0.146 | negtive |
| g__Tropheryma | Thioacetamide | 0.247 | 0.166 | positive |
| g__Brevundimonas | Thioacetamide | −0.201 | 0.260 | negtive |
| g__Aeromonas | Thioacetamide | −0.198 | 0.270 | negtive |
| g__Lactobacillus | Thioacetamide | −0.196 | 0.273 | negtive |
| g__Pseudoalteromonas | Thioacetamide | −0.163 | 0.364 | negtive |
| g__Leifsonia | Thioacetamide | −0.147 | 0.413 | negtive |
| g__Moraxella | Thioacetamide | −0.128 | 0.478 | negtive |
| g__Haemophilus | Thioacetamide | 0.103 | 0.567 | positive |
| g__Porphyromonas | Thioacetamide | 0.101 | 0.577 | positive |
| g__Gardnerella | Thioacetamide | −0.084 | 0.641 | negtive |
| g__Prevotella | Thioacetamide | 0.083 | 0.645 | positive |
| g__Alloprevotella | Thioacetamide | 0.078 | 0.666 | positive |
| g__Neisseria | Thioacetamide | 0.077 | 0.670 | positive |
| g__Veillonella | Thioacetamide | 0.063 | 0.729 | positive |
| g__Streptococcus | Thioacetamide | 0.054 | 0.767 | positive |
| g__Prevotella_7 | Thioacetamide | 0.005 | 0.977 | positive |
| g__Brevundimonas | XMP | 0.422 | 0.015 | positive |
| g__Acinetobacter | XMP | 0.364 | 0.038 | positive |
| g__Aeromonas | XMP | 0.360 | 0.040 | positive |
| g__Pseudoalteromonas | XMP | 0.355 | 0.042 | positive |
| g__Prevotella | XMP | −0.322 | 0.068 | negtive |
| g__Leifsonia | XMP | 0.310 | 0.079 | positive |
| g__Vibrio | XMP | 0.291 | 0.100 | positive |
| g__Hydrogenophaga | XMP | 0.233 | 0.191 | positive |
| g__Lactobacillus | XMP | −0.230 | 0.197 | negtive |
| g__Moraxella | XMP | 0.223 | 0.213 | positive |
| g__Alloprevotella | XMP | −0.175 | 0.331 | negtive |
| g__Haemophilus | XMP | −0.155 | 0.390 | negtive |
| g__Streptococcus | XMP | −0.129 | 0.473 | negtive |
| g__Porphyromonas | XMP | −0.120 | 0.505 | negtive |
| g__Mycoplasma | XMP | 0.074 | 0.681 | positive |
| g__Veillonella | XMP | −0.056 | 0.755 | negtive |
| g__Neisseria | XMP | −0.027 | 0.879 | negtive |
| g__Tropheryma | XMP | 0.020 | 0.914 | positive |
| g__Prevotella_7 | XMP | 0.012 | 0.948 | positive |
| g__Gardnerella | XMP | −0.004 | 0.984 | negtive |
| Pathway | Description | # compounds_ num(dem) | Compounds(dem) | Total | Percent | Rich Factor | Raw_p | -ln(p) | ms2_description |
|---|---|---|---|---|---|---|---|---|---|
| hsa01100 | Metabolic pathways | 27 | C00144;C00164;C00655;C02814;C030 56;C01586;C00793;C00120;C05904;C0 0037;C01112;C00092;C00989;C01005; C00160;C06102;C00159;C00858;C007 19;C03519;C00019;C04501;C05382;C0 0197;C00780;C01879;C00246 | 2974 | 87.097 | 0.009 | 0.016 | 4.146 | Global and overview maps |
| hsa01200 | Carbon metabolism | 6 | C00160;C05382;C00037;C00197;C009 89;C01005 | 114 | 19.355 | 0.053 | 0.000 | 8.992 | Global and overview maps |
| hsa00270 | Cysteine and methionine metabolism | 5 | C00019;C00793;C00197;C0314 5;C01005 | 66 | 16.129 | 0.076 | 0.000 | 9.342 | Amino acid metabolism |
| hsa01230 | Biosynthesis of amino acids | 5 | C00019;C05382;C00197;C0003 7;C01005 | 128 | 16.129 | 0.039 | 0.002 | 6.267 | Global and overview maps |
| hsa00260 | Glycine, serine and threonine metabolism | 4 | C01005;C00197;C00719;C00037 | 48 | 12.903 | 0.083 | 0.000 | 8.005 | Amino acid metabolism |
| hsa02010 | ABC transporters | 4 | C00159;C00719;C00120;C00037 | 138 | 12.903 | 0.029 | 0.016 | 4.145 | Membrane transport |
| hsa01250 | Biosynthesis of nucleotide sugars | 4 | C00159;C04501;C05382;C01112 | 200 | 12.903 | 0.02 | 0.052 | 2.954 | Global and overview maps |
| hsa01240 | Biosynthesis of cofactors | 4 | C00197;C00120;C00037;C00019 | 328 | 12.903 | 0.012 | 0.204 | 1.591 | Global and overview maps |
| hsa05230 | Central carbon metabolism in cancer | 3 | C00092;C00197;C00037 | 37 | 9.677 | 0.081 | 0.002 | 6.128 | Cancer: overview |
| hsa00650 | Butanoate metabolism | 3 | C00164;C00989;C00246 | 47 | 9.677 | 0.064 | 0.004 | 5.441 | Carbohydrate metabolism |
| hsa00630 | Glyoxylate and dicarboxylate metabolism | 3 | C00160;C00197;C00037 | 62 | 9.677 | 0.048 | 0.009 | 4.666 | Carbohydrate metabolism |
| hsa00230 | Purine metabolism | 3 | C00144;C00037;C00655 | 99 | 9.677 | 0.030 | 0.033 | 3.421 | Nucleotide metabolism |
| hsa04721 | Synaptic vesicle cycle | 2 | C00780;C00037 | 12 | 6.452 | 0.167 | 0.003 | 5.769 | Nervous system |
| hsa04973 | Carbohydrate digestion and absorption | 2 | C00092;C00246 | 27 | 6.452 | 0.074 | 0.016 | 4.165 | Digestive system |
| hsa04742 | Taste transduction | 2 | C00144;C00780 | 32 | 6.452 | 0.063 | 0.021 | 3.841 | Sensory system |
| hsa00030 | Pentose phosphate pathway | 2 | C00197;C05382 | 35 | 6.452 | 0.057 | 0.025 | 3.672 | Carbohydrate metabolism |
| hsa00480 | Glutathione metabolism | 2 | C01879;C00037 | 38 | 6.452 | 0.053 | 0.030 | 3.519 | Metabolism of other amino acids |
| hsa04974 | Protein digestion and absorption | 2 | C00037;C00246 | 47 | 6.452 | 0.043 | 0.044 | 3.128 | Digestive system |
| hsa00120 | Primary bile acid biosynthesis | 2 | C15519;C00037 | 47 | 6.452 | 0.043 | 0.044 | 3.128 | Lipid metabolism |
| hsa00310 | Lysine degradation | 2 | C00164;C00037 | 50 | 6.452 | 0.04 | 0.049 | 3.016 | Amino acid metabolism |
| hsa04080 | Neuroactive ligand-receptor interaction | 2 | C00780;C00037 | 52 | 6.452 | 0.038 | 0.053 | 2.946 | Signaling molecules and interaction |
| hsa00970 | Aminoacyl-tRNA biosynthesis | 2 | C01005;C00037 | 52 | 6.452 | 0.038 | 0.053 | 2.946 | Translation |
| hsa05208 | Chemical carcinogenesis - reactive oxygen species | 2 | C02814;C00019 | 57 | 6.452 | 0.035 | 0.062 | 2.782 | Cancer: overview |
| hsa00470 | D-Amino acid metabolism | 2 | C00793;C00037 | 67 | 6.452 | 0.030 | 0.082 | 2.500 | Metabolism of other amino acids |
| hsa00520 | Amino sugar and nucleotide sugar metabolism | 2 | C00159;C04501 | 118 | 6.452 | 0.017 | 0.206 | 1.581 | Carbohydrate metabolism |
| hsa04142 | Lysosome | 1 | C00159 | 4 | 3.226 | 0.25 | 0.028 | 3.565 | Transport and catabolism |
| hsa04740 | Olfactory transduction | 1 | C00144 | 8 | 3.226 | 0.125 | 0.056 | 2.886 | Sensory system |
| hsa04744 | Phototransduction | 1 | C00144 | 8 | 3.226 | 0.125 | 0.056 | 2.886 | Sensory system |
| hsa04022 | cGMP-PKG signaling pathway | 1 | C00144 | 10 | 3.226 | 0.1 | 0.069 | 2.669 | Signal transduction |
| hsa04122 | Sulfur relay system | 1 | C00019 | 11 | 3.226 | 0.091 | 0.076 | 2.578 | Folding, sorting and degradation |
| hsa04540 | Gap junction | 1 | C00780 | 11 | 3.226 | 0.091 | 0.076 | 2.578 | Cellular community – eukaryotes |
| hsa04625 | C-type lectin receptor signaling pathway | 1 | C00159 | 11 | 3.226 | 0.091 | 0.076 | 2.578 | Immune system |
| hsa04917 | Prolactin signaling pathway | 1 | C00092 | 11 | 3.226 | 0.091 | 0.076 | 2.578 | Endocrine system |
| hsa04911 | Insulin secretion | 1 | C00092 | 12 | 3.226 | 0.083 | 0.083 | 2.494 | Endocrine system |
| hsa01523 | Antifolate resistance | 1 | C00144 | 17 | 3.226 | 0.059 | 0.115 | 2.163 | Drug resistance: antineoplastic |
| hsa04931 | Insulin resistance | 1 | C00092 | 19 | 3.226 | 0.053 | 0.128 | 2.058 | Endocrine and metabolic disease |
| hsa04918 | Thyroid hormone synthesis | 1 | C00092 | 21 | 3.226 | 0.048 | 0.140 | 1.965 | Endocrine system |
| hsa04024 | cAMP signaling pathway | 1 | C00780 | 25 | 3.226 | 0.04 | 0.165 | 1.804 | Signal transduction |
| hsa04922 | Glucagon signaling pathway | 1 | C00197 | 26 | 3.226 | 0.038 | 0.171 | 1.768 | Endocrine system |
| hsa00780 | Biotin metabolism | 1 | C00120 | 29 | 3.226 | 0.034 | 0.188 | 1.669 | Metabolism of cofactors and vitamins |
| hsa04978 | Mineral absorption | 1 | C00037 | 29 | 3.226 | 0.034 | 0.188 | 1.669 | Digestive system |
| hsa05207 | Chemical carcinogenesis - receptor activation | 1 | C00780 | 29 | 3.226 | 0.034 | 0.188 | 1.669 | Cancer: overview |
| hsa00010 | Glycolysis / Gluconeogenesis | 1 | C00197 | 31 | 3.226 | 0.032 | 0.200 | 1.609 | Carbohydrate metabolism |
| hsa00730 | Thiamine metabolism | 1 | C00037 | 31 | 3.226 | 0.032 | 0.200 | 1.609 | Metabolism of cofactors and vitamins |
| hsa00620 | Pyruvate metabolism | 1 | C03248 | 32 | 3.226 | 0.031 | 0.206 | 1.581 | Carbohydrate metabolism |
| hsa04750 | Inflammatory mediator regulation of TRP channels | 1 | C00780 | 35 | 3.226 | 0.029 | 0.223 | 1.501 | Sensory system |
| hsa00500 | Starch and sucrose metabolism | 1 | C00092 | 37 | 3.226 | 0.027 | 0.234 | 1.453 | Carbohydrate metabolism |
| hsa00561 | Glycerolipid metabolism | 1 | C00197 | 38 | 3.226 | 0.026 | 0.239 | 1.429 | Lipid metabolism |
| hsa04977 | Vitamin digestion and absorption | 1 | C00120 | 39 | 3.226 | 0.026 | 0.245 | 1.407 | Digestive system |
| hsa05415 | Diabetic cardiomyopathy | 1 | C00092 | 39 | 3.226 | 0.026 | 0.245 | 1.407 | Cardiovascular disease |
| hsa04726 | Serotonergic synapse | 1 | C00780 | 42 | 3.226 | 0.024 | 0.261 | 1.342 | Nervous system |
| hsa00280 | Valine, leucine and isoleucine degradation | 1 | C00164 | 42 | 3.226 | 0.024 | 0.261 | 1.342 | Amino acid metabolism |
| hsa00052 | Galactose metabolism | 1 | C00159 | 46 | 3.226 | 0.022 | 0.282 | 1.265 | Carbohydrate metabolism |
| hsa00562 | Inositol phosphate metabolism | 1 | C00092 | 47 | 3.226 | 0.021 | 0.287 | 1.247 | Carbohydrate metabolism |
| hsa00360 | Phenylalanine metabolism | 1 | C03519 | 49 | 3.226 | 0.020 | 0.298 | 1.211 | Amino acid metabolism |
| hsa00051 | Fructose and mannose metabolism | 1 | C00159 | 54 | 3.226 | 0.019 | 0.323 | 1.131 | Carbohydrate metabolism |
| hsa00760 | Nicotinate and nicotinamide metabolism | 1 | C03056 | 55 | 3.226 | 0.018 | 0.328 | 1.116 | Metabolism of cofactors and vitamins |
| hsa00440 | Phosphonate and phosphinate metabolism | 1 | C00037 | 56 | 3.226 | 0.018 | 0.333 | 1.101 | Metabolism of other amino acids |
| hsa00330 | Arginine and proline metabolism | 1 | C00019 | 72 | 3.226 | 0.014 | 0.406 | 0.902 | Amino acid metabolism |
| hsa00350 | Tyrosine metabolism | 1 | C00164 | 78 | 3.226 | 0.013 | 0.431 | 0.841 | Amino acid metabolism |
| hsa05204 | Chemical carcinogenesis - DNA adducts | 1 | C14704 | 81 | 3.226 | 0.012 | 0.444 | 0.813 | Cancer: overview |
| hsa00524 | Neomycin, kanamycin and gentamicin biosynthesis | 1 | C00092 | 81 | 3.226 | 0.012 | 0.444 | 0.813 | Biosynthesis of other secondary metabolites |
| hsa00380 | Tryptophan metabolism | 1 | C00780 | 83 | 3.226 | 0.012 | 0.452 | 0.795 | Amino acid metabolism |
| hsa04976 | Bile secretion | 1 | C00780 | 97 | 3.226 | 0.010 | 0.505 | 0.683 | Digestive system |
| hsa00860 | Porphyrin metabolism | 1 | C00037 | 148 | 3.226 | 0.007 | 0.660 | 0.415 | Metabolism of cofactors and vitamins |