
Download

ORIGINAL ARTICLE
Utility of mast cell P815 for evaluating FcεRI-dependent and FcεRI-independent allergic effects
Ryosuke Misawaa, Kazuhito Takemotob, Masaki Ijib, Luo Haoa, Akari Koresawab, Hiroyuki Watanabeb*
aGraduate School of Human Life Sciences, University of Kochi, Kochi City, Kochi, Japan
bFaculty of Nutrition, University of Kochi, Kochi city, Kochi, Japan
Abstract
Introduction and Objectives: The degranulation and release of inflammatory mediators mediated by the high-affinity immunoglobulin E (IgE) receptor (FcεRI) on mast cells in response to allergen contact is the driving force of anaphylaxis. This study shows that P815 cells, which were previously thought not to express FcεRI, cause a reaction similar to FcεRI-mediated degranulation in the presence of antigen and IgE.
Materials and Methods: The kinetics of degranulation were evaluated by comparing P815 cells with FcεRI-expressing a rat basophilic leukemia (RBL-2H3) (that typically indicates the specific clone or subline within that cell line) cell lines. Degranulation activity was measured using the release rate of β-hexosaminidase as an indicator. P815 cells showed significant degranulation when compound 48/80 or anti-dinitrophenyl (DNP)-IgE antibody and DNP-human serum albumin (HSA) antigen were added simultaneously. Gene expression analysis confirmed the expression of each FcεRI subunit—specifically, the γ subunit expressed markedly. Moreover, the expression of the phosphorylation enzymes Lyn, spleen tyrosine kinase (Syk), Fyn, and Bruton tyrosine kinase (Btk), which are involved in degranulation, was upregulated.
Results: FcεRI has three subunits: α, β, and γ. P815 cells do not express FcεRI because they have messenger ribonucleic acid (mRNA) for the γ subunit but not for the α and β subunits. However, P815 expressed each subunit protein (α, β, and γ), as detected in the western blotting analysis of cell extracts in the presence of DNP-HSA antigen and anti-DNP-IgE.
Conclusion: These results suggest that P815 may cause degranulation via FcεRI. Therefore, P815 is considered to be a cell model that can evaluate both FcεRI-mediated and FcεRI-independent degranulation reactions in response to allergens.
Key words: anaphylaxis, G protein-coupled receptor, mast cell degranulation, signal transduction, mast cell
*Corresponding author: Hiroyuki Watanabe, Faculty of Nutrition, University of Kochi, Kochi city, Kochi, Japan. Email address: [email protected]
Received 13 November 2024; Accepted 3 March 2025; Available online 1 June 2025
Copyright: Misawa R, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Anaphylaxis is an acute life-threatening reaction that develops rapidly after contact with a triggering substance. Classically, anaphylaxis is an allergen-driven process that induces specific immunoglobulin E (IgE) and activates mast cells and basophils via IgE receptor cross-linking. IgE antibodies bind to type I high-affinity IgE receptors (FcεRIs) on mast cells and basophils, sensitizing them to being triggered in a subsequent encounter with the cognate allergen.1
The FcεRI consists of three subunits: α, β, and γ. FcεRI is expressed as the αβγ2 tetramer and αγ2 trimer in humans, whereas it is expressed on the surface of mast cells and basophils as an αβγ2 tetramer in rodents. Among the three subunits, the α chain binds directly to IgE and is an essential molecule in IgE-mediated allergic reactions. However, the β and γ subunits are involved in cell signal transduction.2,3
P815 is a mast cell line isolated from a mouse mastocytoma.4 The messenger ribonucleic acid (mRNA) for the γ subunit in the cells comprises the IgE receptor; however, the lack of mRNA for the receptor’s α and β subunits hinders FcεRI expression.5 The transfection of complementary deoxyribonucleic acids (cDNAs) for the α and β subunits results in the formation of stable clones that can synthesize abundant α, β, and γ mRNA to express normal FcεRI.
Most studies have focused on the FcεRI to examine mast cell degranulation, as it requires sensitization with IgE and activation by secondary exposure to agonists;6 however, there are also mast cell receptors that do not require IgE for activation known as non-IgE receptors.7 For the non-IgE receptor response, only one stimulation is required for activation through the non-IgE receptors. Examples of non-IgE receptors on mast cells include toll-like receptors (e.g., TLR4), G protein-coupled receptors (e.g., C3aR), and alarmin receptors (e.g., interleukin (IL)-1R).8–10
The classical mast cell agonist 48/80 (C48/80), an exogenous polymer of N-methyl-p-methoxyphenethylamine condensed with formaldehyde, has been used to study non-IgE stimulation. This agonist activates mast cells through the mas-related G protein-coupled receptor X1/X2 (MRGPRX1/X2) in humans, B2 analog (MRGPRB2) in mice, and B3 analog (MRGPRB3) in rats.11–13
These findings suggest that mast cells are activated by various pathways independent of FcεRI and serve a protective function.
Although some studies indicate that degranulation responses are activated independently of the IgE pathway in P815,14,15 there are no fundamental studies on the degranulation responsiveness of the IgE and non-IgE pathways in P815 cells. Therefore, in this study, we demonstrate the degranulation kinetics of P815 in light of the degranulation of rat basophilic leukemia (RBL-2H3) (that typically indicates the specific clone or subline within that cell line) cell lines,16 which express the FcεRI, and proposed the existence of an activation pathway mediated by FcεRI in P815 cells. Furthermore, we propose that the degranulation reaction caused by P815 could be used as a model to improve the accuracy of research into anaphylaxis caused by mast cells.
Materials and Methods
Cells
Mouse mastocytoma P815 and RBL-2H3 cell lines were obtained from the RIKEN BioResource Center (Ibaraki, Japan).
Cell culture
P815 cells were cultured in RPMI 1640 (Nacalai Tesque, Inc., Kyoto, Japan) containing inactivated 10% fetal bovine serum (Sigma-Aldrich Japan, Tokyo, Japan) and 1% antibiotic-antifungal mixture (Nacalai Tesque, Inc.). RBL-2H3 cells were cultured in low-glucose Dulbecco’s modified Eagle’s medium (Nacalai Tesque, Inc.) at 37°C in a humidified 5% carbon dioxide (CO2) atmosphere.
Basic degranulation activity assay
Degranulation activity was compared based on the β-hexosaminidase (Hex) release rate from RBL-2H3 and P815.17
RBL-2H3 or P815 cell suspension (100 µL) adjusted to 2.5 × 105 cells/mL, and then the well was seeded into the 96-well microplates and cultured for 24 hours until 90% confluency. The cells were sensitized with culture medium containing rat anti-dinitrophenyl (DNP)-IgE antibody (final concentration 0.45 µg/mL; Yamasa Corporation, Tokyo, Japan) or mouse anti-DNP-IgE antibody (final concentration 0.45 µg/mL; Yamasa Corporation) for 24 hours. The cells were subsequently degranulated for 2 hours and after that, the culture medium was aspirated. Then, 120 µL of MT buffer was added to the cells and heated at 37°C for 10 minutes, followed by treatment with 20 µL of 10 µg/mL of DNP that serves as an antigen (Invitrogen, Inc., Carlsbad, CA). The reaction was quenched on ice for 10 minutes and 50 µL of the supernatant was warmed at 37°C for 5 minutes before incubation with 3.3 mmol/L of p-nitrophenyl-2-acetoamido-2-deoxy-β-D glucopyranoside (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) in 100 µL of 0.1 mol/L citrate buffer (pH 4.5) in a 96-well plate. Hex was then added and incubated at 37°C for 25 minutes. The reaction was stopped by the addition of 100 µL of 2.0 mol/L glycine buffer (pH 10.4) and the absorbance was measured at 405 nm using a plate reader (iMark, Bio-Rad Laboratories, Inc., Tokyo, Japan).
Cells remaining in the 96-well microplate were homogenized using an ultrasonic device (Cross Ultrasonic Homogenizer Elestein NP-035SD; Elecom Science Co., Chiba, Japan) to obtain intracellular fluid; then the 50 µL of the intracellular fluid was transferred to a 96-well plate and heated at 37°C for 5 minutes. Next, 3.3 mmol/L p-nitrophenyl-2-acetoamido-2-deoxy-β-D glucopyranoside (FUJIFILM Wako Pure Chemical Corporation) in 100 µL of 0.1 mol/L citrate buffer (pH 4.5) was added. Hex was then added, and the plate was incubated at 37°C for 25 minutes. Subsequently, the reaction was stopped by adding 100 µL of 2.0 mol/L glycine buffer (pH 10.4) and the absorbance was measured at 405 nm using a plate reader.
The Hex release rate was determined using the following formula: Hex release rate (%) = amount released/(amount released + amount remaining in cells) × 100.
The degranulation test was compared under the following four conditions, including positive and negative controls:
-
Degranulation by addition of C48/80
The Hex reaction was performed by adding only C48/80 without anti-DNP-IgE antibody and DNP-HSA antigen.
-
Degranulation by the addition of DNP-HSA
The Hex reaction was performed by adding only DNP-HSA antigen without adding an anti-DNP-IgE antibody.
-
Degranulation with anti-DNP-IgE antibodies
The Hex reaction was performed by adding only the anti-DNP-IgE antibody and no DNP-HSA antigen.
-
Degranulation by addition of anti-DNP-IgE antibody and DNP-HSA antigen.
Hex reactions were performed without the addition of anti-DNP-IgE antibody and DNP-HSA antigen.
Real-time polymerase chain reaction
P815 cells were subjected to degranulation tests. The mRNA was prepared using Agencourt RNAdvance Tissue (Beckman Coulter, Inc., Brea, CA, USA) and stored at 80°C until use. Complementary DNA was prepared by reverse transcription using PrimeScript™ RT reagent kit (Takara Bio Inc., Shiga, Japan).
Reverse transcription polymerase chain reaction (RT-PCR) was performed using Probe qPCR Mix (Takara Bio Inc.). The expression of α, β, and γ subunits of FcεRI, MRGPRB2, Lyn, Syk, Fyn, and Btk gene in each cell was quantitated using a TaqMan® Probe (α subunit: Mm00438867_m1, β subunit: Mm00442778_m1, γ subunit: Mm02343757_m1, MRGPRB2: Mm01956240_s1, Lyn: Mm01217488_m1, Syk: Mm01333032_m1, Fyn: Mm01304022_m1, and Btk: Mm00442712_m1; Thermo Fisher Scientific, Tokyo, Japan). Gene expression was normalized to actin expression as an internal standard. The Ct (threshold cycle) values of each gene relative to that of actin in the same cell were calculated as relative expression of the target gene.
The expression of the nonreceptor phosphatases, Lyn, Syk, Fyn, and Btk involved in degranulation was done similarly. The results were normalized to actin as an internal standard and calculated as relative values.
Western blot analysis
For the degranulation tests, 2.0 × 105 cells were collected. Then, 30 µg of total protein were subjected to Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) using 10% or 15% polyacrylamide gels containing 0.1% SDS. FcεRIα and β subunits were separated by electrophoresis (AE-6530, ATTO Corporation, Tokyo, Japan). SDS was removed using the ProteinAssay CBB Clean Up kit (Nacalai Tesque, Inc.), followed by the Coomassie brilliant blue staining18 (Protein Assay CBB Solution, Nacalai Tesque, Inc.). Subsequently, the protein was transferred to a polyvinylidene difluoride (PVDF) membrane (WSE-4051, ATTO Corporation, Tokyo, Japan) by semidry blotting (AE-6677, ATTO Corporation). The membranes were blocked with 7.5% bovine serum albumin (FUJIFILM Wako Pure Chemical Corporation) and incubated with primary antibodies (Fc Epsilon Rα: Fc Epsilon RI Alpha Rabbit Polyclonal FcεRβ: Anti-MS4A2 Polyclonal Antibody, Bioss Inc., MA, USA; FcεRγ: Anti-FCER1G, Rabbit, GeneTex, International Corporation, Hsinchu City, Taiwan), followed by horseradish peroxidase-conjugated secondary antibody (Vectastain® Elite ABC-HRP kit, Peroxidase [Rabbit IgG] Kit, Vector Laboratories, Inc., CA, USA). Finally, the membranes were developed using horseradish peroxidase substrate (ImmPACT® DAB Substrate kit, Peroxidase [HRP], Vector Laboratories, Inc.) for color development.
Ethics approval
These cells were transferred from RIKEN (Rikagaku Kenkyūsho). There were no usage-specific regulations, and additionally there were no special legal regulations or restrictions for other matters. We hereby declare that this report is in line with the guidelines of this journal.
Statistical analysis
The values obtained are expressed as the mean ± standard deviation for each group. Statistical analysis was performed using the statistical software Excel Statistics (Social Information Service Co. Ltd, Tokyo, Japan).
The results of degranulation were compared using the Steel method with a reagent concentration of zero used as a control. The results of gene expression were compared using the Steel method with a group without DNP-HSA antigen and anti-DNP-IgE antibody used as a control and other groups. The significance level was set at < 0.05 or < 0.01.
Results
Degranulation responsiveness
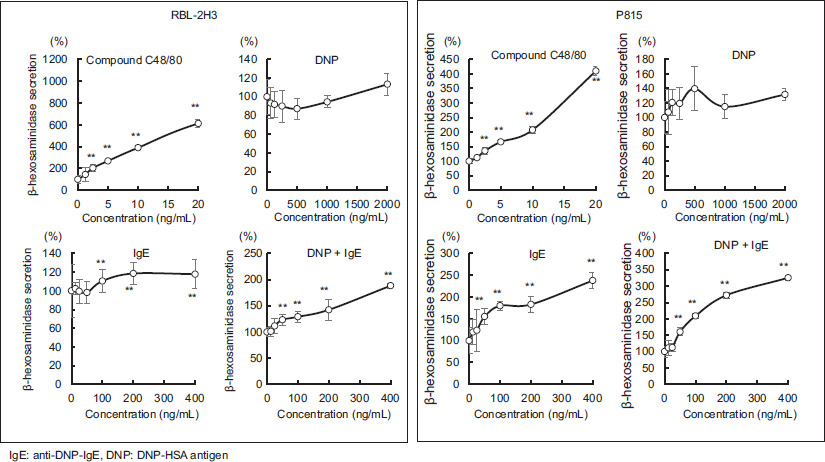
The responsiveness of RBL-2H3 and P815 cells to the degranulation assessment is shown in Figure 1, compared with and without the addition of each substance. The results are presented as the percentage of Hex released outside the cells (%).
Figure 1 Changes in the release rate of hexosaminidase after addition of various reagents to RBL-2H3 or P815. Values are mean ± standard deviation (n=4). Significant difference compared using the Steel method, with a reagent concentration of zero used as a control. **: p<0.01
In the RBL-2H3 cells, the Hex release rate increased in a C48/80 concentration-dependent manner; however, the addition of DNP-HSA antigen alone did not alter the Hex release rate. The addition of anti-DNP-IgE antibodies increased the Hex release rate only slightly in a dose-dependent manner, whereas the simultaneous addition of anti-DNP-IgE antibody and DNP-HSA antigen markedly increased the Hex release rate.
In P815 cells, the Hex release rate increased with the addition of C48/80; however, it was not as high as that in RBL-2H3. Adding DNP-HSA antigen alone did not significantly alter the Hex release rate, similar to that in RBL-2H3. In contrast, when anti-DNP-IgE antibodies were added alone, the rate of Hex release increased in proportion to the amount of anti-DNP-IgE antibodies and was higher than that of RBL-2H3. Furthermore, the simultaneous addition of anti-DNP-IgE antibodies and DNP-HSA antigens significantly increased the Hex release rate in a dose-dependent manner compared with that in RBL-2H3 cells.
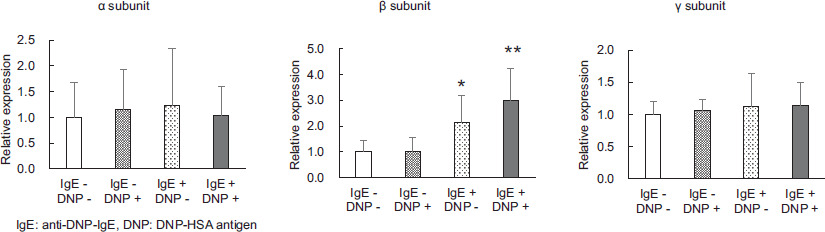
Expression of the FcεRI constitutive subunits
The intensity of the expression of each subunit comprising FcεRI in cells subjected to degranulation tests is shown in Figure 2. Intensity in the basic degranulation test was compared when both the DNP-HSA antigen and anti-DNP-IgE antibody were not added. The α and γ subunits were expressed at similar levels with and without the addition of DNP-HSA antigen and anti-DNP-IgE antibody, whereas the β subunit was upregulated by the DNP-HSA antigen alone (P < 0.05) and with the simultaneous addition of DNP-HSA antigen and anti-DNP-IgE antibody (P < 0.01).
Figure 2 The effect of adding antigens and/or antibodies to P815 on the expression levels of the various subunit genes that comprise FcεRI. Values are mean ± standard deviation (n=9). Significant difference compared using the Steel method, with a group without DNP-HSA antigen and anti-DNP-IgE antibody used as a control, and other groups. *: p<0.05, **: p<0.01.
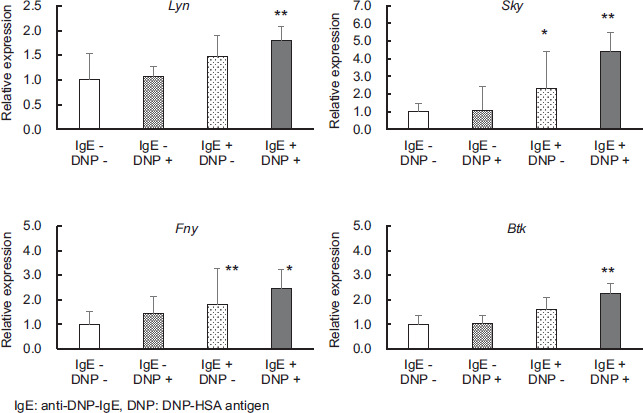
Expression of phosphatase genes involved in degranulation
The expression level of the various phosphorylated enzymes in cells subjected to the degranulation tests is shown in Figure 3. Relative expression was compared when both DNP-HSA antigen and anti-DNP-IgE antibody were not added to the cells. Under degranulation conditions, the expression of Lyn and Btk were significantly increased when both DNP-HSA antigen and anti-DNP-IgE antibody were added compared with their expression when both DNP-HSA antigen and anti-DNP-IgE antibody were not added (P < 0.01). The expression of Syk and Fyn was significantly increased when the anti-DNP-IgE antibody was added alone (Syk: P < 0.05, Fyn: P < 0.01) and they were significantly increased when both the antigen NP-HSA and the anti-DNP-IgE antibody were added (Syk: P < 0.01, Fyn: P < 0.05).
Figure 3 Effects of adding antigens and/or antibodies to P815 on the expression levels of various phosphorylated protein genes involving FcεRI. Values are mean ± standard deviation (n=9). Significant difference compared using the Steel method, with a group without DNP-HSA antigen and anti-DNP-IgE antibody used as a control, and other groups. *: p<0.05, **: p<0.01.
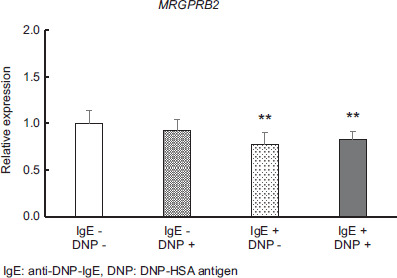
Gene expression of MRGPRB2 receptors
The expression of the MRGPRB2 receptor in cells subjected to the degranulation tests is shown in Figure 4, which was compared when both DNP-HSA antigen and anti-DNP-IgE antibody were not added. The MRGPRB2 receptor was downregulated with the addition of anti-DNP-IgE antibody alone (P < 0.01) and by the addition of both DNP-HSA antigen and anti-DNP-IgE antibody (P < 0.01).
Figure 4 Effect of adding antigens and/or antibodies to P815 on the expression level of the MRGPRB2 gene. Values are mean ± standard deviation (n=9). Significant difference compared using the Steel method, with a group without DNP-HSA antigen and anti-DNP-IgE antibody used as a control, and other groups. *: p<0.05; **: p<0.01.
Expression of genes related to degranulation
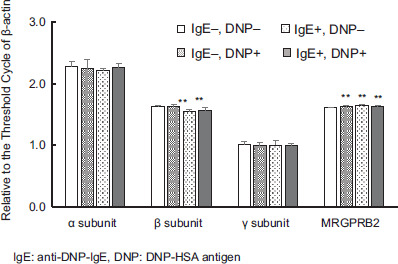
The Ct values for each subunit comprising FcεRI in the cells subjected to the degranulation tests relative to the Ct value of β-actin are shown in Figure 5, and a comparison of the intensity of the basic degranulation test when both DNP-HSA antigen and anti-DNP-IgE antibodies were not added is also shown. The relative Ct values for the γ subunit were the smallest, followed by the β subunit, whereas the α subunit showed the largest values. The relative Ct values of the β subunit decreased with the addition of DNP-HSA antigen alone (P < 0.01) and with the simultaneous addition of DNP-HSA antigen and anti-DNP-IgE antibody (P < 0.01).
Figure 5 Intensity of expression of various genes by addition of antigen and/or antibody to P815. Values are for Threshold Cycle of each gene relative to Threshold Cycle of β-actin, mean ± standard deviation (n=9). Significant difference compared using the Steel method, with a group without DNP-HSA antigen and anti-DNP-IgE antibody used as a control, and other groups. **: p<0.01.
Figure 6 Detection by Western blotting of the various subunits constituting FcεRI after addition of antigen and/or antibody to P815.
Mouse liver tumor cells (Hepa 1-6) and mouse skeletal muscle tumor cells (C2C12) are lineages other than rat basophilic mast cells and mouse mast cells. These cell lines showed no expression of the individual subunits constituting FcεRI (data not shown), whereas MRGPRB2 exhibited a Ct value similar to that of the β subunit of FcεRI.
Western blot analysis
Each subunit expressed in the cells subjected to the degranulation tests is shown in Figure 5, the α subunit with a molecular weight of 50.7 kDa is shown in Figure 5A, the β subunit with a molecular weight of 32.4 kDa is shown in Figure 5B, and the γ subunit with a molecular weight of 14.0 kDa is shown in Figure 5C.
Discussion
Mast cells are the major effector cells of allergic reactions and play a critical role during the development of allergenic diseases. These cells secrete histamine and various inflammatory and immunomodulatory substances produced during anaphylactic reactions. In this study, the degranulation response in P815 mast cells was confirmed in comparison to the RBL-2H3 cells.
Reportedly, monomeric IgE may enhance mast cell activity without the cross-linking of FcεRI by IgE-specific allergens or auto-reactive immunoglobulin G (IgG) anti-IgE antibodies.19 Herein, degranulation occurred with anti-DNP-IgE antibodies alone, suggesting that P815 may be responding to monomeric IgE. Conversely, when anti-DNP-IgE antibody and DNP-HSA antigen were added simultaneously, degranulation was induced with a strength 1.4–1.8 times of that induced by IgE alone; this suggests that P815 induces degranulation via FcεRI by simultaneously adding IgE and antigen.
The addition of DNP-HSA antigen and anti-DNP-IgE caused marked degranulation in P815 cells, similar to that in RBL-2H3 cells. Thus, the factors that cause degranulation were the addition of the coexistence of DNP-HSA antigen and anti-DNP-IgE. This suggests that P815 cells, like RBL-2H3 cells, trigger FcεRI receptor-mediated degranulation.
Additionally, this study shows that the addition of C48/80 caused non-IgE-mediated degranulation in RBL-2H3 and P815, as reported previously.20,21
The expression of the three subunits of FcεRI was observed under the four degranulation conditions in P815 cells. The expression of the γ subunit was the highest, and interestingly, the expression of the β subunit was increased by the addition of anti-DNP-IgE. Based on the Ct value of actin as a reference, the α subunit had a lower expression level compared with that of MRGPRB2, whereas the β subunit had a similar level. The expression of α, β, and γ subunit proteins was also observed in P815 cell extracts in the presence of DNP-HSA antigen and anti-DNP-IgE under the four degranulation conditions.
The IgE binding to the FcεRI receptor on mast cells resulted in aggregation, which activated the tyrosine kinases Lyn, Fyn, Hck, and Fgr of the Src family bound to the β chain followed by the β and γ chains’ immunoreceptor tyrosine-based activation motif, which phosphorylates tyrosine residues. The tyrosine-phosphorylated γ chain binds Syk, which is phosphorylated and activated by Lyn and Syk. The tyrosine kinase phosphorylates adaptor molecules, such as linker for activation of T cells (LAT), SLP (stromal lymphopoietin)-76, tyrosine kinase Btk, and Vav guanine nucleotide exchange factor (VAV), leading to mast cell activation.17,18 This phosphorylation triggers degranulation via calcium efflux through the activation of phospholipase Cγ1/2 and phosphatidylinositol-3-kinase, as reported in previous studies.24,25
In this study, increased expression of the Lyn, Fyn, Syk, and Btk genes was observed in the presence of DNP-HSA antigen and anti-DNP-IgE under the four degranulation conditions using P815 cells. Thus, it is assumed that degranulation is triggered through calcium release by phospholipase Cγ1/2. These findings suggest that P815 cells trigger FcεRI-mediated degranulation.
Both RBL-2H3 and P815 cells show degranulation following the addition of C48/80, which suggests that the degranulation is not mediated by the FcεRI receptor but by receptors of the MRGPR system (RBL-2H3: MRGPRA2, P815: MRGPRB2). No differences were detected in the expression of the MRGPRB2 gene in the mouse MRGPR system receptors in cells subjected to either degranulation test.
G proteins can be divided into three types—stimulatory G protein (Gs), inhibitory G protein (Gi), and G protein that activates phospholipase C (Gq)—based on the differences in Gα. MRGPRX2 and MRGPRB2 signaling is regulated by Gα members, followed by phospholipase C (PLC) activation, and then phosphatidylinositol 4,5-diphosphate (PIP2) to diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IP3). Subsequently, IP3 activates the calcium channels in the endoplasmic reticulum, inducing degranulation.26 Thus, MRGPRB2 may play a role in P815 cell signal degranulation through this pathway.
In a primary bone marrow-derived mast cell-macrophage co-culture system, P815 cells enhanced IL-4-mediated uptake and replication of the Francisella tularensis live vaccine strain (LVS) within the macrophages.27 The co-cultures of primary mast cells suggested that P815 cells can regulate LVS replication within macrophages by producing IL-4 and mast cell activation by non-FcεR-driven signaling when treated with mast cell-activating peptides.
5-Hydroxymethylfurfural (5-HMF) is produced by the Maillard or caramelization reaction of reducing sugars at high temperatures, as 5-HMF exerts an immunosensitization reaction when injected into mice. Li et al. used P815 cells to elucidate the potential immunotoxicity caused by anaphylaxis-like reactions mediated by 5-HMF-containing preparations and related mechanistic analyses of non-FcεRI-mediated anaphylaxis-like reactions.28
In this study, we found that although the expression of the α subunit which comprises FcεRI is relatively small in P815 cells, the β- and γ subunit genes are relatively intensely expressed in FcεRI. Researchers may use the results of both non-FcεRI-mediated and FcεRI-mediated degranulation separately, depending on how they determine the index of degranulation and the environment.
In earlier studies, we used the FcεRI-dependent degranulation reaction of P815 cells to evaluate the degranulation in mouse-derived cells using MCP-5 and PT-1829,30; however, P815 can induce significant degranulation. Furthermore, type I allergy assays can be conducted seamlessly using the mouse-derived cell line P815 and mice.
Finally, we also emphasized the differentiation between basophils and mast cells that share several characteristics—they have basophilic granules in the cytoplasm, their cell surfaces express FcεRIs, and they release chemical mediators such as histamine.31 However, basophils are the least common granulocytes, accounting for less than 1% of the circulating white blood cells. Because of this similarity and their minority status, basophils had often been erroneously considered minor relatives or blood-circulating precursors of tissue-resident mast cells; hence long been neglected or underestimated in immunological studies.32 However, basophils and mast cells differ in several aspects: basophils normally circulate in the blood, whereas mast cells are found in peripheral tissues, and basophils have a much shorter lifespan than mast cells. Furthermore, the gene expression profiles of basophils and mast cells differ markedly in mice and humans,33,34 suggesting a unique role of basophils that is different from that of mast cells.35 Additionally, basophil activation tests are often used to diagnose food allergies; however, the role of basophils in the development of food allergies remains unclear.36–39
Therefore, the method of measuring degranulation using P815 mast cells to evaluate anti-allergic activity is considered an important evaluation method that emphasizes the characteristics of mast cells, which are the site of basic immediate allergic reactions.
Conclusion
In this study, we confirm that P815 cells, which have been assumed not to express the FcεRI, show activation by monomeric IgE when IgE antibody is added alone and show degranulation via the IgE receptor when antigen and IgE antibody are added simultaneously. In allergy research, we show that P815 cells can be used to assess FcεRI-dependent and nondependent degranulation responses. The results of this study may contribute to improving the accuracy of anaphylaxis research using mast cells.
Author Contributions
All author’s contributed equally to this article.
Conflict of Interest
The authors declare no potential conflicts of interest with respect to research, authorship, and/or publication of this article.
Funding
None.
REFERENCES
1 Vitte J, Vibhushan S, Bratti M, Montero-Hernandez JE, Blank U. Allergy, anaphylaxis, and nonallergic hypersensitivity: IgE, mast cells, and beyond. Med Princ Pract. 2022;31(6):501–15. 10.1159/000527481
2 Blank U, Ra C, Miller L, White K, Metzger H, Kinet JP. Complete structure and expression in transfected cells of high affinity IgE receptor. Nature. 1989;337 (6203):187–9. 10.1038/337187a0
3 Ra C, Jouvin MH, Kinet JP. Complete structure of the mouse mast cell receptor for IgE (Fc epsilon RI) and surface expression of chimeric receptors (rat-mouse-human) on transfected cells. J Biol Chem. 1989;264(26):15323–7. 10.1016/S0021-9258(19)84829-9
4 Dunn TB, Potter M. A transplantable mast-cell neoplasm in the mouse. J Natl Cancer Inst. 1957;18(4):587–601.
5 Miller L, Alber G, Varin-Blank N, Ludowyke R, Metzger H. Transmembrane signaling in P815 mastocytoma cells by transfected IgE receptors. J Biol Chem. 1990;265(21):12444–53. 10.1016/S0021-9258(19)38366-8
6 Nagata Y, Suzuki R. FcεRI. A Master regulator of mast cell functions. Cells. 2022;11(4):622. 10.3390/cells11040622
7 Yu Y, Blokhuis BR, Garssen J, Redegeld FA. Non-IgE mediated mast cell activation. Eur J Pharmacol. 2016;778:33–43. 10.1016/j.ejphar.2015.07.017
8 Nilsson G, Johnell, Hammer CH, Tiffany HL, Nilsson K, Metcalfe DD, et al. C3a and C5a are chemotaxins for human mast cells and act through distinct receptors via a pertussis toxin-sensitive signal transduction pathway. J Immunol. 1996;157(4):1693–8.
9 Kirshenbaum AS, Swindle E, Kulka M, Wu Y, Metcalfe DD. Effect of lipopolysaccharide (LPS) and peptidoglycan (PGN) on human mast cell numbers, cytokine production, and protease composition. BMC Immunol. 2008;9:45. 10.1186/1471-2172-9-45
10 Hültner L, Kölsch S, Stassen M, Kaspers U, Kremer J-P, Mailhammer R, et al. In activated mast cells, IL-1 upregulates the production of several Th2-related cytokines including IL-9. J Immunol. 2000;164(11):5556–63. 10.4049/jimmunol.164.11.5556
11 Irman-Florjanc T, Erjavec F. Compound 48/80 and substance P induced release of histamine and serotonin from rat peritoneal mast cells. Agents Act. 1983;13(2-3):138–41. 10.1007/BF01967317
12 Kashem SW, Subramanian H, Collington SJ, Magotti P, Lambris JD, Ali H. G protein coupled receptor specificity for C3a and compound 48/80-induced degranulation in human mast cells: Roles of mas-related genes MrgX1 and MrgX2. Eur J Pharmacol. 2011;668(1-2):299–304. 10.1016/j.ejphar.2011.06.027
13 Zylka MJ, Dong X, Southwell AL, Anderson DJ. Atypical expansion in mice of the sensory neuron-specific Mrg G protein-coupled receptor family. Proc Natl Acad Sci USA. 2003;100(17):10043–8. 10.1073/pnas.1732949100
14 Wang N, Liu R, Liu Y, Zhang R, He L. Sinomenine potentiates P815 cell degranulation via upregulation of Ca2+ mobilization through the Lyn/PLCγ/IP3R pathway. Int J Immunopathol Pharmacol. 2016;29(4):676–83. 10.1177/0394632015621768
15 Hu X, Wen Y, Liu S, Luo J, Tan X, Li Z, et al. Evaluation of the anaphylactoid potential of andrographis paniculata extracts using the popliteal lymph node assay and P815 cell degranulation in vitro. J Transl Med. 2015;13:121. 10.1186/s12967-015-0478-0
16 Péterfy H, Tóth G, Pecht I, Erdei A. C3a-derived peptide binds to the type I FcepsilonR and inhibits proximal-coupling signal processes and cytokine secretion by mast cells. Int Immunol. 2008;20(10):1239–45. 10.1093/intimm/dxn083
17 Matsuda H, Nakamura S, Yoshikawa M. Degranulation inhibitors from medicinal plants in antigen-stimulated rat basophilic leukemia (RBL-2H3) cells. Chem Pharm Bull. 2016;64(2):96–103. 10.1248/cpb.c15-00781
18 Goldring JPD. Measuring protein concentration with absorbance, Lowry, Bradford Coomassie Blue, or the Smith bicinchoninic acid assay before electrophoresis. Methods Mol Biol. 2019;1855:31–9. 10.1007/978-1-4939-8793-1_3
19 Kalesnikoff J, Huber M, Lam V, Damen JE, Zhang J, Siraganian RP, et al. Monomeric IgE stimulates signaling pathways in mast cells that lead to cytokine production and cell survival. Immunity. 2001;14(6):801–11. 10.1016/s1074-7613(01)00159-5
20 Wang J, Liu J, Yang Y, Sun G, Yang D, Yin S, et al. Inhibitory effect of phellodendrine on C48/80-induced allergic reaction in vitro and in vivo. Int Immunopharmacol. 2024;134:112256. 10.1016/j.intimp.2024.112256
21 Zhang X, Yao H, Qian Q, Li N, Jin W, Qian Y. Cerebral mast cells participate in postoperative cognitive dysfunction by promoting astrocyte activation. Cell Physiol Biochem. 2016;40(1-2):104–16. 10.1159/000452528
22 Bugajev V, Bambousková M, Dráberová L, Dráber P. What precedes the initial tyrosine phosphorylation of the high affinity IgE receptor in antigen-activated mast cell? FEBS Lett. 2010;584(2-4):4949–55. 10.1016/j.febslet.2010.08.045
23 Gilfillan AM, Rivera J. The tyrosine kinase network regulating mast cell activation. Immunol Rev. 2009;228(1):149–69. 10.1111/j.1600-065X.2008.00742.x
24 Kageyama-Yahara N, Wang X, Katagiri T, Wang P, Yamamoto T, Tominaga M, et al. Suppression of phospholipase Cγ1 phosphorylation by cinnamaldehyde inhibits antigen-induced extracellular calcium influx and degranulation in mucosal mast cells. Biochem Biophys Res Commun. 2011;416(3-4):283–8. 10.1016/j.bbrc.2011.11.014
25 Ito N, Yokomizo T, Sasaki T, Kurosu H, Penninger J, Kanaho Y, et al. Requirement of phosphatidylinositol 3-kinase activation and calcium influx for leukotriene B4-induced enzyme release. J Biol Chem. 2002;277(47):44898–904. 10.1074/jbc.M208051200
26 Wang Z, Franke K, Bal G, Li Z, Zuberbier T, Babina M. MRGPRX2-mediated degranulation of human skin mast cells requires the operation of Gαi, Gαq, Ca++ channels, ERK1/2 and PI3K—Interconnection between early and late signaling. Cells. 2022;11(6):953. 10.3390/cells11060953
27 Thathiah P, Sanapala S, Rodriguez AR, Yu JJ, Murthy AK, Guentzel MN, et al. Non-FcεR bearing mast cells secrete sufficient interleukin-4 to control Francisella tularensis replication within macrophages. Cytokine. 2011;55(2):211–20. 10.1016/j.cyto.2011.04.009
28 Li E, Lin N, Hao R, Fan X, Lin L, Hu G, et al. 5-HMF induces anaphylactoid reactions in vivo and in vitro. Toxicol Rep. 2020;7:1402–11. 10.1016/j.toxrep.2020.10.010
29 Tashiro M, Kawakami Y, Abe R, Han W, Hata D, Sugie K, et al. Increased secretion of TNF-alpha by costimulation of mast cells via CD28 and Fc epsilon RI. J Immunol. 1997;158(5):2382–9.
30 Fukamachi H, Yamada N, Miura T, Kato T, Ishikawa M, Gulbins E, et al. Identification of a protein, SPY75, with repetitive helix-turn-helix motifs and an SH3 domain as a major substrate for protein tyrosine kinase(s) activated by Fc epsilon RI cross-linking. J Immunol. 1994;152(2):642–52.
31 Li Y, Leung PSC, Gershwin ME, Song J. New mechanistic advances in fcεri-mast cell-mediated allergic signaling. Clin Rev Allergy Immunol. 2022;63(3):431–46. 10.1007/s12016-022-08955-9
32 Karasuyama H, Miyake K, Yoshikawa S, Kawano Y, Yamanishi Y. How do basophils contribute to Th2 cell differentiation and allergic responses? Int Immunol. 2018;30(9):391–6. 10.1093/intimm/dxy026
33 Dwyer DF, Barrett NA, Austen KF. Immunological genome project C. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat Immunol. 2016;17(7):878–87. 10.1038/ni.3445
34 Motakis E, Guhl S, Ishizu Y, Itoh M, Kawaji H, de Hoon M, et al. Redefinition of the human mast cell transcriptome by Deep-CAGE sequencing. Blood. 2014;123(17):e58–67. 10.1182/blood-2013-02-483792
35 Miyake K, Ito J, Karasuyama H. Role of basophils in a broad spectrum of disorders. Front Immunol. 2022;13:902494. 10.3389/fimmu.2022.902494
36 Hussain M, Borcard L, Walsh KP, Pena Rodriguez M, Mueller C, Kim BS, et al. Basophil-derived IL-4 promotes epicutaneous antigen sensitization concomitant with the development of food allergy. J Allergy Clin Immunol. 2018;141(1):223–34. 10.1016/j.jaci.2017.02.035
37 Muto T, Fukuoka A, Kabashima K, Ziegler SF, Nakanishi K, Matsushita K, et al. The role of basophils and proallergic cytokines, thymic SLP and IL-33, in cutaneously sensitized food allergy. Int Immunol. 2014;26(10):539–49. 10.1093/intimm/dxu058
38 Noti M, Kim BS, Siracusa MC, Rak GD, Kubo M, Moghaddam AE, et al. Exposure to food allergens through inflamed skin promotes intestinal food allergy through the thymic stromal lymphopoietin-basophil axis. J Allergy Clin Immunol. 2014;133(5):1390–9. 10.1016/j.jaci.2014.01.021
39 Miyake K, Shibata S, Yoshikawa S, Karasuyama H. Basophils and their effector molecules in allergic disorders. Allergy. 2021;76(6):1693–706. 10.1111/all.14662